
Ribonucleic acid (RNA) plays a complex and central role in cellular regulation, catalysis, and signal transduction. Visualizing RNA in isolation and in the natural cellular environment is an important means of elucidating its functions. The ability to image populations of RNA enables monitoring of cellular transcriptional activity, tracking the migration and dynamics of various transcripts, and analyzing RNA-rich subcellular compartments and biomolecular aggregates (such as nucleoli, P bodies, and stress granules). However, despite decades of effort, imaging populations of RNA in vitro and in cells and tissues remains a fundamental challenge in the field of biomolecular visualization chemistry.
Recently, RNA labeling and imaging strategies have primarily focused on specific transcripts rather than broad-spectrum imaging. Significant advances include aptamer-based systems, such as Spinach and Mango aptamers, which bind dyes through engineered RNA motifs and trigger a significant increase in emission intensity. Although these systems are suitable for specific engineered transcripts, the aptamer modules cannot be applied to populations of RNA in natural cellular environments. Molecular beacons, as another major RNA detection and imaging strategy, are short-chain deoxyribonucleic acid (DNA) hairpin oligonucleotides labeled with fluorophores and quenchers. These modified DNA produce fluorescence upon hybridization with target RNA through sequence complementarity and have been applied in areas such as real-time polymerase chain reaction (PCR), but their action is sequence-specific rather than broad-spectrum, and they can produce significant background fluorescence in cellular applications. Other hybridization probes (such as locked nucleic acids (LNA) and peptide nucleic acids (PNA) have higher affinity and nuclease resistance but still rely on sequence-specific targeting and are often limited to fixed cell imaging. Enzyme-assisted RNA labeling strategies (such as tRNA guanylyltransferase (TGT) mediated labeling, DNAzyme labeling, or CRISPR–Cas13 guided labeling) provide covalent or modular labeling schemes for RNA, but they require sequence engineering of RNA or only target specific transcripts, thus not suitable for general RNA populations.
Recently, bioorthogonal metabolic incorporation techniques for nucleosides have achieved RNA labeling across the entire transcriptome, thus breaking the sequence limitation. In this strategy, modified nucleosides are taken up by cells, phosphorylated by kinases, and subsequently incorporated into newly synthesized RNA. For example, alkyne-containing 5-ethynyluridine (5 EU) and N6-propynyladenosine can be further functionalized through click chemistry. However, this method requires fixed cells for fluorescence labeling and washing off excess dye, making it unsuitable for widespread applications in live-cell RNA imaging and not applicable for gel visualization of natural RNA.
There is an urgent need to develop strategies based on the intrinsic chemical properties of RNA (rather than sequence) that can achieve direct and sequence-independent high-selectivity labeling. To this end, small molecule fluorescent dyes that bind to RNA through non-covalent interactions are being developed for broad-spectrum imaging of this biopolymer, with the advantage of structural tunability. These push-pull fluorescent groups based on aromatic cations non-covalently bind to RNA through electrostatic attraction and intercalation, with emission wavelengths tunable over a wide range, and some are cell membrane permeable, making them suitable for real-time imaging and gel staining. The earliest dye used for gel RNA imaging is the intercalator ethidium bromide (EtBr), which preferentially stains DNA, but can also stain RNA. Recently developed SYBR series commercial fluorescent dyes (such as SYBR Green and SYBR Gold) bind to nucleic acids through electrostatic interactions, intercalation, or groove binding, exhibiting higher sensitivity. SYBR Green II can stain both RNA and single-stranded DNA (ssDNA), while SYBR Gold preferentially stains double-stranded DNA (dsDNA), but can also weakly stain ssDNA and RNA. The commercial dye SYTO RNASelect is designed for selective staining of live-cell RNA, but recent data show its limited cell permeability and lower selectivity for RNA compared to DNA (only 1.6 times). Additionally, other fluorophore designs aim to enhance sensitivity, selectivity, and imaging performance.QIUD-2 (a styryl dicationic dye) exhibits superior photostability and biocompatibility compared to SYTO RNASelect, enabling live-cell RNA imaging, but requires washing steps prior to imaging. Push-pull cationic dyes containing methylpyridine or methylquinoline enhance fluorescence over 100 times upon binding to RNA, staining nucleolar and mitochondrial RNA, but the latter may be difficult to distinguish due to the structural similarity of mitochondrial organelle dyes. Probes based on naphthalimide exhibit a 32-fold fluorescence turn-on upon binding to rRNA, with rapid cellular uptake and excellent photostability. Indolizine-containing styryl cationic dyes may provide the highest signal enhancement (490 times) among known RNA dyes and enable subcellular visualization through fluorescence lifetime imaging (FLIM). Finally, PYQU serves as a multifunctional probe capable of dual-channel detection of RNA and sulfur dioxide (SO2), achieving endogenous RNA visualization in cultured cells and zebrafish through deep red fluorescence, although its selectivity and fluorescence turn-on data have not been reported. Despite some probes showing improvements in sensitivity and biocompatibility, they all rely on non-covalent binding mechanisms based on intercalation and electrostatic interactions, which can increase off-target background fluorescence due to negatively charged substances and hydrophobic environments, making it difficult to distinguish RNA from DNA. Notably, cationic aromatic fluorophores structurally similar to previously reported RNA dyes are also widely used for mitochondrial labeling, revealing background and specificity issues caused by such low-specificity interactions. Similarly, common commercial RNA dyes exhibit extremely low or no selectivity for RNA compared to DNA. These limitations highlight the necessity of utilizing the unique physicochemical properties of RNA to achieve high selectivity and stable signal generation in labeling strategies.
Recent studies have revealed key chemical features of RNA as a biopolymer: the presence of the 2′-hydroxyl (2′–OH) group in ribose. This functional group is absent in DNA, constituting an inherent chemical selectivity control point that provides a chemical basis for selective targeting of RNA. The pKa value of the 2′–OH in RNA (approximately 12.5) is lower than that of reference alcohols (ethanol pKa=15.9) and exhibits abnormally high reactivity towards electrophiles. Functionalized electrophiles that can efficiently react with 2′–OH include acyl, sulfonyl, and aryl electrophiles. These reactions can occur efficiently under mild aqueous conditions, with reaction rates and specificities comparable to those of protein thiol modifications in some cases. Covalent reagents developed for targeting 2′–OH modifications have been applied in structural probing (such as SHAPE), labeling, RNA protection and delivery, RNA purification, and interaction analysis, while maintaining the natural structure of RNA. Acyl derivatives on RNA can be designed to be reversible through chemical triggering or catalyst design, releasing unmodified RNA for subsequent analysis. Reagents targeting 2′–OH can possess tunable reactivity, solubility, and cell compatibility, promoting emerging applications from whole transcriptome structural mapping to live-cell RNA imaging and the development of RNA targeting chemical tools.
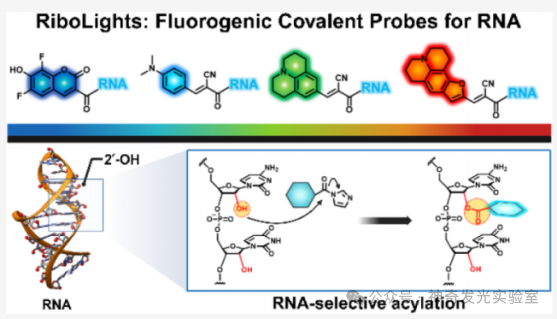
To address the limitations of existing broad-spectrum RNA labeling and staining strategies, this article develops a molecular approach utilizing the reactivity of 2′–OH. A class of previously unexplored fluorescent generating acyl probes that covalently react with RNA is reported. These tunable reagents (named RiboLight (RL) fluorophores) can achieve sequence-independent RNA labeling while possessing high selectivity and significant signal enhancement. Notably, RL fluorophores are non-cationic, thus not relying on non-specific electrostatic attraction and target interactions. Each RL fluorophore is designed to emit in a specific spectral window, enabling multicolor imaging and ratio analysis. By integrating fluorescence generation activation with covalent bond formation in a single step, the RL platform overcomes key drawbacks of non-covalent RNA fluorophores, such as low specificity, transient binding, limited signal enhancement, and the need for washing steps. This probe is compatible with both gel and cellular RNA imaging and is effective across various RNA sequences and structural environments. Importantly, the absence of 2′–OH in DNA ensures that RL fluorophores have very low cross-reactivity with DNA, achieving a high level of RNA:DNA selectivity. Finally, it is found that these fluorophores can be removed from the target under mild conditions, restoring unmodified RNA for subsequent analysis. These features collectively establish RL fluorophores as powerful chemical tools for in vitro and intracellular RNA imaging and analysis.
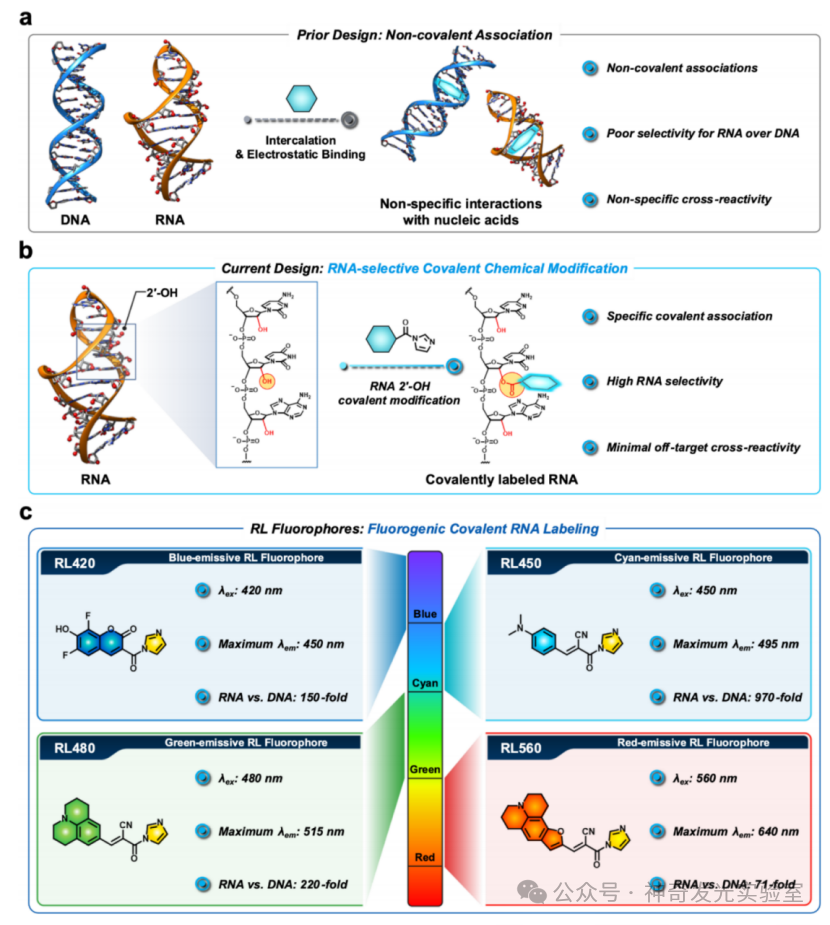
Figure 1 Development of RNA-selective fluorescent covalent acyl probes (RiboLight, RL)
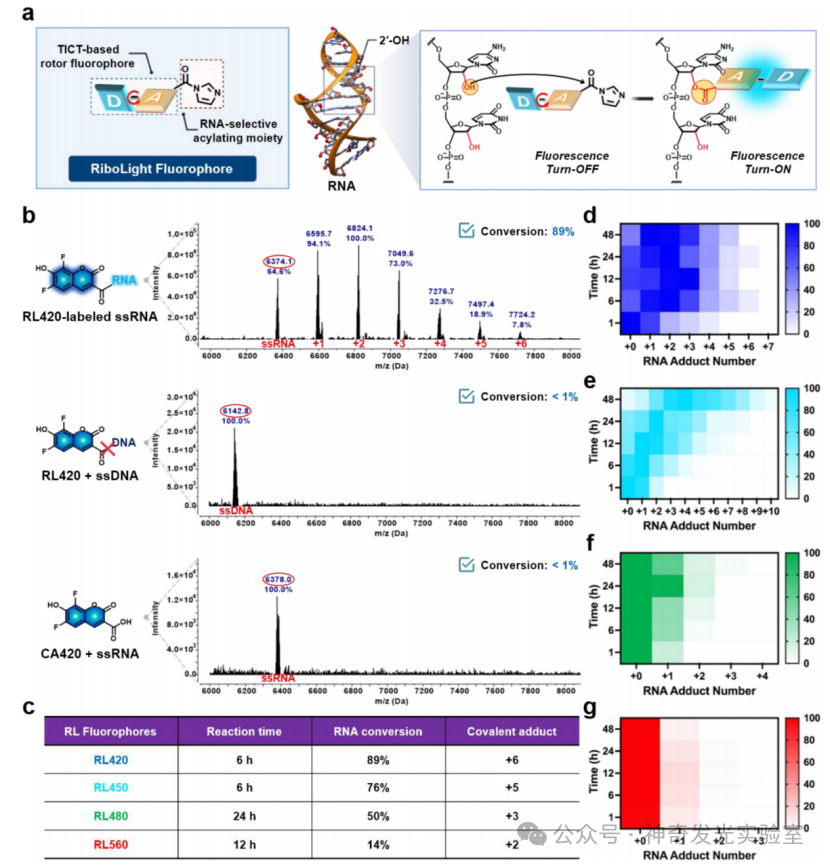
Figure 2 Covalent labeling of RNA through RL fluorophores achieving 2′-hydroxyl selective acylation
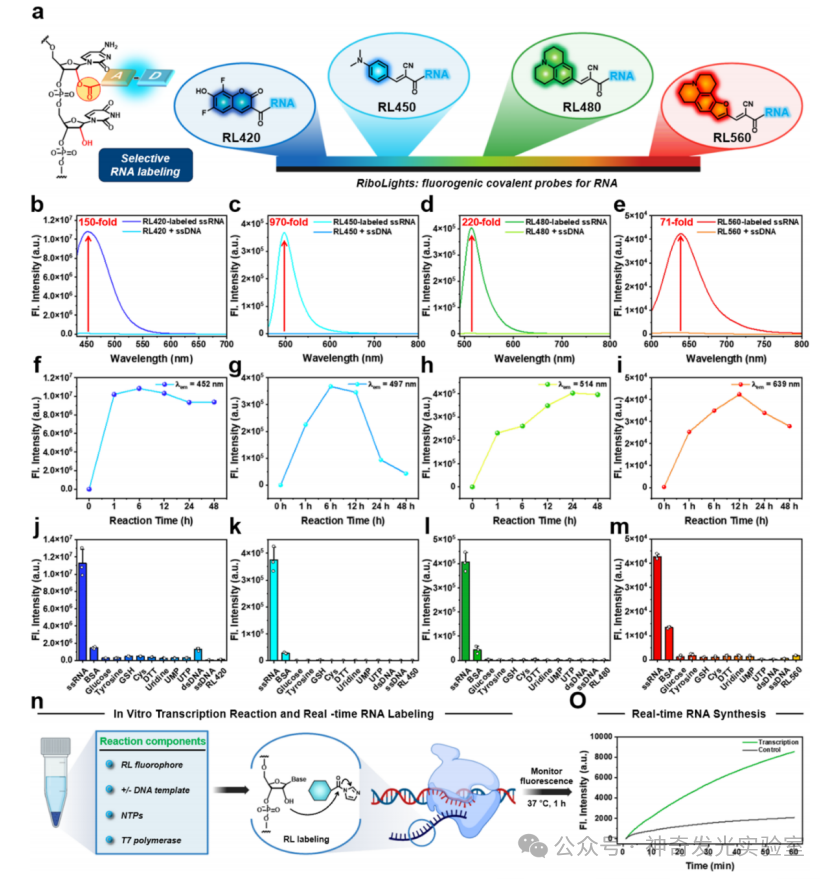
Figure 3 Multicolor fluorescence RNA labeling achieved through selective 2′-hydroxyl acylation with RL fluorophores
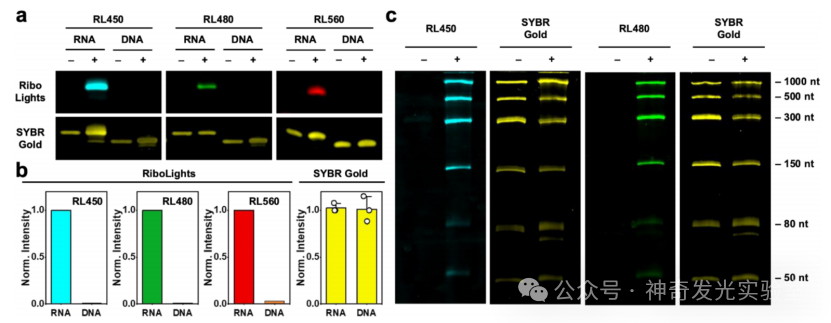
Figure 4 Selective gel imaging of RNA based on RL fluorophores
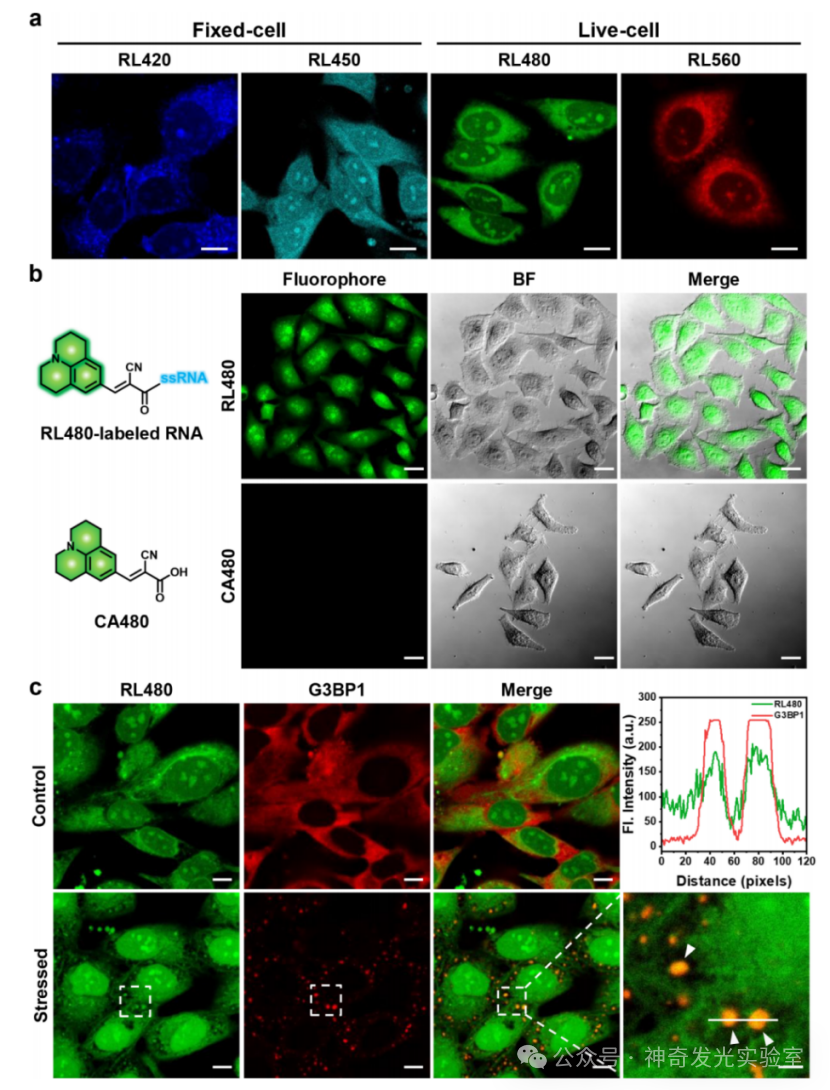
Figure 5 Cellular imaging of RNA using RL fluorophores
[Reference Details] Jinwoo Shin, Moon Jung Kim, and Eric T. Kool. Fluorogenic Covalent Probes for RNA. J. Am. Chem. Soc., 2025, https://doi.org/10.1021/jacs.5c14938