
Click the blue text to follow us!
Source: Journal of Cancer Prevention and Treatment, 2020, Volume 47 Issue 12
Qiu Jinmei1, Gong Juan1, Xie Qingchi1, Xie Zhizhong1,2
Affiliations:1. 421001 Hengyang, Key Laboratory of Tumor Microenvironment Response Drug Research, Nanhua University;2. 421001 Hengyang, Hunan Provincial Collaborative Innovation Center for New Drug Research on Molecular Targets, Nanhua University
Funding: Hunan Province Graduate Research Innovation Project 2019 (CX2019 0748); Hunan Province College Student Innovation and Entrepreneurship Training Program Project 2019 (2324)
|
2020,47(12):980-985. DOI:10.3971/j.issn.1000-8578.2020.20.0279 |
【Abstract】Abnormal energy metabolism is one of the characteristic pathological changes of tumors. Studies have shown that tumor cells preferentially utilize glycolysis to generate lactate and a small amount of ATP even in the presence of oxygen, aided by lactate dehydrogenase A (LDHA), which is known as the Warburg effect. Excess lactate within cells is transported outside via the monocarboxylate transporter 4 (MCT4) on the cell membrane, leading to a decrease in the pH of the tumor cell microenvironment. Meanwhile, lactate in the tumor microenvironment can be transported by MCT1 to adjacent relatively oxygen-rich tumor cells and converted back to pyruvate for oxidative phosphorylation under the action of LDHB. The important role of LDH in the reprogramming of tumor energy metabolism makes it a new target for antitumor therapy. This article reviews the relationship between LDH and tumor cell energy metabolism, focusing on the role of LDH in tumor development and the progress of research on antitumor drugs targeting LDH.
【Keywords】Lactate;Lactate Dehydrogenase;Warburg Effect;Pyruvate Metabolism
0
Introduction
Cells take up glucose through glucose transporters and convert it to pyruvate through glycolysis, which can then follow two possible metabolic pathways: one is that when oxygen supply is sufficient, pyruvate enters the mitochondria and undergoes aerobic oxidation, ultimately converting to CO2 and H2O, producing a large amount of adenosine triphosphate (ATP); the other is when oxygen supply is insufficient or cannot be fully utilized, pyruvate does not enter the mitochondria but is reduced to lactate by lactate dehydrogenase A (LDHA) in the cytoplasm, simultaneously producing a small amount of ATP. In the 1920s, the famous German biochemist Warburg discovered[1] that tumor cells primarily metabolize glucose through the latter pathway even in aerobic conditions to provide energy, a metabolic mode known as the “Warburg effect,” characterized by high glucose consumption, low ATP synthesis, and high lactate production. For a long time, lactate produced by this pathway was regarded as a metabolic waste and overlooked. However, recent studies have shown that the lactate generated by this mechanism can be taken up by adjacent tumor cells that are relatively deficient in glucose supply and, with the assistance of lactate dehydrogenase B (LDHB), can be converted back to pyruvate for ATP synthesis, providing energy necessary for rapid tumor cell proliferation. Blocking lactate production and utilization through drugs is expected to become another effective means for targeted tumor therapy. This article focuses on the relationship between LDH and tumor cell energy metabolism, as well as the progress of research on antitumor drugs targeting LDH, aiming to provide new insights into the pathogenesis of tumors and the development of antitumor drugs.
1
Transmembrane Transport of Lactate in Tumor Cells
When tumor cells choose aerobic glycolysis as their main metabolic mode, a large amount of lactate will be generated within the cells, and if the lactate level is too high, it will have direct cytotoxic effects on the cells themselves. Therefore, tumor cells must transport excess lactate out of the cells. Studies have confirmed that monocarboxylate transporters (MCTs) are the main transporters for exporting lactate from inside the cells[2].
MCTs are a class of transmembrane proteins widely distributed on mammalian cell membranes, consisting of 14 members, all encoded by the SLC16A gene family, which are highly conserved across various species. However, only four (MCT1, MCT2, MCT3, and MCT4) have been confirmed to participate in the H+ coupled transport of monocarboxylic acids (including lactate, pyruvate, and ketone bodies), among which MCT1 is present in almost all tissues, especially in the heart and fibroblasts. MCT2 is mainly found in the liver, kidneys, and nerve cells. MCT3 was initially expressed in chicken and rat retinal pigment epithelial cells, localized to the basal membrane of RPE. MCT4 is commonly found in cells that heavily rely on glycolysis, such as white muscle fibers, leukocytes, and tumor cells[2–3].
The role of different MCTs in lactate efflux varies under different tissues and conditions. For tumor cells in a relatively hypoxic environment, MCT4 is considered the most important subtype for lactate efflux. Studies have confirmed that MCT4 is upregulated in various tumors, including triple-negative breast cancer, clear cell renal carcinoma, pancreatic cancer, and non-small cell lung cancer, while blocking MCT4 leads to increased acidity within the cells, causing “acid stress” and ultimately inducing tumor cell apoptosis[4–6].
Under physiological conditions, the lactate concentration in blood or tissues is approximately 1.5-3.0 mmol[7], but in tumors, due to MCT4-mediated lactate efflux, the lactate concentration in blood or tissues can rise to 10-30 mmol[8–9]. This acidic tumor microenvironment is also one of the characteristic pathological changes of tumors, while the lactate in the tumor interstitium can serve as an important carbon source for adjacent tumor cells in relatively oxygen-rich areas, participating in the reprogramming of tumor energy metabolism. The uptake of lactate by these adjacent tumor cells is primarily mediated by MCT1. Studies have shown that in tumor cells with a high rate of aerobic glycolysis, inhibiting MCT1 using MCT1 inhibitors or gene silencing techniques can suppress the uptake of lactate and ketone bodies, significantly reducing mitochondrial aerobic phosphorylation in tumor cells and inhibiting tumor cell growth[10–11].
2
The Role and Application of Lactate Dehydrogenase in Tumors
Lactate dehydrogenase (LDH) is a zinc-containing metalloprotein with a relative molecular weight of 135-140 kD, which is a glycolytic enzyme and one of the important metabolic enzymes in the tumor microenvironment. LDH is a tetramer composed of two different subunits, namely LDHA and LDHB. They can assemble into five different tetramers (isoenzymes): LDH1 consists of four LDHB subunits; LDH2 contains three LDHB and one LDHA subunit; LDH3 contains two LDHB and two LDHA subunits; LDH4 has one LDHB and three LDHA subunits; and LDH5 consists of four LDHA subunits. In nature, due to differences in substrate selectivity, there are two different isoforms of LDH, L(-)- and D(+)-LDH. In invertebrates, protists, and bacteria, D(+)-LDH is primarily present, which is a 2-hydroxy acid dehydrogenase and belongs to the FAD-binding oxidoreductase family; while in vertebrate tissues (including humans), L(-)-LDH is predominant, which is an L-specific NAD-dependent dehydrogenase. The LDH discussed in this article mainly refers to the latter.
LDH is widely distributed in the body, almost present in the cytoplasm of all tissue cells, especially in high concentrations in the liver, kidneys, and myocardium. Due to its relatively large molecular weight, it is mainly distributed in the cytoplasm, and only when pathological conditions arise, such as damage to myocardial or liver cells, does LDH spill from the cytoplasm, leading to abnormal elevation of LDH levels in plasma. Therefore, plasma LDH levels are commonly used as important indicators for clinical detection of heart and liver function.
LDH mediates the bidirectional conversion between pyruvate and lactate and plays an important role in regulating energy metabolism between tumor cells. Many tumor patients exhibit elevated levels of LDH in their blood or cancerous secretions, and this elevation of LDH levels has even been used for the diagnosis and prognostic analysis of related cancers[12–13].
2.1 LDHA and Lactate Metabolism and Its Role in Tumors
The energy metabolism of tumor cells is highly plastic, allowing them to choose specific energy metabolic pathways based on their living environment. Generally, tumor cells proliferate at a much faster rate than normal cells, significantly increasing their energy demand. Tumor cells located in hypoxic areas drive glucose uptake by promoting the expression of glucose transporters (GLUTs) on the cell membrane. However, for these tumor cells, glucose is not oxidized to pyruvate and enters the mitochondria for oxidative phosphorylation (OXPHOS); instead, it is broken down into lactate under the action of LDHA, producing NAD+ and a small amount of ATP (1 molecule of glucose ultimately produces 2 molecules of lactate and 2 ATP), demonstrating a metabolic mode of “high glucose uptake and low ATP generation,” which is the well-known “Warburg effect.” Since this mode produces only a small amount of ATP (under normal circumstances, 1 molecule of glucose can produce up to 38 ATP via OXPHOS), it has been regarded by scientists as a “wasteful” energy metabolic pattern. Warburg discovered early on that for tumor cells, this “wasteful metabolic pattern” is still adopted for glycolysis even under aerobic conditions, and the reason why tumor cells prefer this metabolic mode has remained a mystery for the scientific community[14].
Warburg’s early view suggested that the loss of mitochondrial function in cancer cells and the low function of the respiratory chain prevent pyruvate from entering the mitochondria for effective oxidative phosphorylation. However, current studies have confirmed that there are indeed cases where mitochondrial function in tumor cells changes, but often, tumor cells exhibiting the “Warburg effect” do not show significant damage to mitochondrial function[15], indicating that there are other reasons at play. Although the efficiency of ATP production in the Warburg effect is low, its energy supply speed far exceeds that of normal glucose OXPHOS, helping tumor cells meet the energy demands for rapid proliferation[16]; some also believe that this metabolic pathway leads to increased lactate production within tumor cells, which, when expelled into the tumor interstitium, acidifies the tumor microenvironment, promoting tumor cell proliferation, invasion, migration, drug resistance, and immune evasion[17]; another viewpoint suggests that the lactate produced by this pathway can be taken up by adjacent tumor cells with energy supply obstacles, converted back to pyruvate, and reused for OXPHOS, facilitating the redistribution of energy substances for tumor cells to adapt to different environments[18]; other studies have shown[19] that not only ATP but also rapid tumor cell proliferation requires more substances like proteins and nucleic acids, and tumor cells can utilize intermediate products provided by this glycolysis (such as lactate as a carbon source and NADH as an essential substance for fatty acid synthesis) to promote de novo synthesis of sugars, amino acids, and fats, achieving “metabolic reprogramming” for nucleic acids, lipids, and proteins.
The conversion of pyruvate to lactate in the cytoplasm during glycolysis in the Warburg effect relies on LDHA, and studies have confirmed that LDHA is highly expressed in various tumors and plays a crucial role in tumor development: Cui et al.[20] reported that the transcription factor FOXM1 can promote the development of pancreatic cancer by upregulating LDHA expression; Jiang et al.[21] confirmed that the upregulation of LDHA expression induced by the FOXM1-LDHA signaling pathway plays a significant role in gastric cancer development; Su et al.[22] demonstrated that a histone demethylase JMJD2A can upregulate LDHA expression and promote the proliferation, invasion, and metastasis of nasopharyngeal carcinoma cells; while Jiang et al.[23] found that increased LDHA expression can promote epithelial-mesenchymal transition in bladder cancer cells, accelerating the progression of bladder cancer.
2.2 LDHB and Lactate Metabolism and Its Role in Tumors
In contrast to LDHA, LDHB mainly catalyzes the conversion of lactate to pyruvate and NADH. Recent studies[24] have found that not all tumor cells exhibit low levels of OXPHOS; some tumor cells even show high levels of OXPHOS. Based on this, the “lactate shuttle theory” has been proposed, which posits that for tumor cells located far from blood vessels or in relatively hypoxic areas (hypoxic cancer cells), tumor cells will upregulate GLUTs, take up large amounts of glucose, which is then converted to lactate (including ketone bodies and fatty acids) via glycolysis. These substances are expelled into the tumor interstitium through corresponding mechanisms and can be taken up by adjacent (or oxygen-rich) tumor cells (oxygenated cancer cells) as the main energy supply source, converted back to pyruvate under the action of LDHB, and further enter the mitochondria for OXPHOS to provide ATP, as seen in Figure 1. This energy metabolic mode helps tumor cells allocate energy substances more effectively, ensuring that marginal (or relatively hypoxic) tumor cells can take up more glucose to meet the energy demands of rapid proliferation, while the lactate they produce can guarantee the energy supply for tumor cells in oxygen-rich areas.
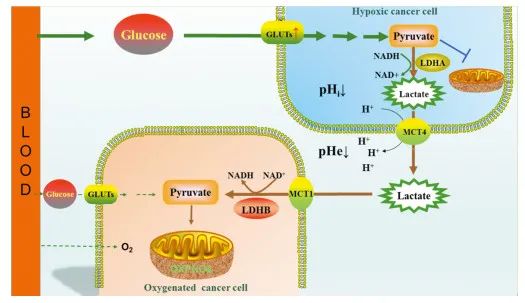 |
| Figure 1“Lactate Shuttle Mechanism” Diagram |
In 2011, Martinez-Outschoorn et al.[25] discovered that when co-culturing tumor cells with fibroblasts, the mitochondrial activity of tumor cells significantly increased, while glucose uptake and GLUT1 expression correspondingly decreased. At the same time, the secretion of ROS (reactive oxygen species, ROS) such as H2O2 significantly increased, which, after diffusing to adjacent fibroblasts, caused an increase in oxidative stress levels within the latter, leading to a significant decrease in mitochondrial activity in fibroblasts, while glucose uptake increased, enhancing aerobic glycolysis. This experimental result not only confirmed the role of the tumor microenvironment in tumor development but also first proposed the role of ROS in tumor cell energy metabolism. Subsequently, the “reverse Warburg effect” theory gradually developed on this basis, which states that the aerobic glycolysis level of stromal cells in the tumor microenvironment is enhanced, and the lactate and ketone bodies produced are taken up by tumor cells, promoting OXPHOS in tumor cells.
Generally, the expression level of LDHA in tumor cells is significantly increased, but the expression of LDHB varies among different tumor cells due to the differences in tumor types. Studies have confirmed that LDHB expression is upregulated in cervical cancer (such as SiHa cells)[26], lung cancer cell lines (such as non-small cell lung cancer NCI-H226, KRAS mutant lung adenocarcinoma NCI-H2122, NCI-H1573, etc.)[27], osteosarcoma (such as MG63, U2OS, and SAOS cells)[28], oral squamous cell carcinoma (such as HN12, Cal27, and KB cells)[29], polyomavirus negative Merkel cell lines[30], and some corresponding clinical cases. LDHB is downregulated or not expressed in liver cancer cells SNU-423, SNU-449, and MHCC97-H[31], colon cancer cells RKO and HCT116[32], gastric cancer cells KATOⅢ[33], and highly metastatic prostate cancer cells LNCaP-LN3[34].
Moreover, even within the same tumor type, the expression of LDHB can vary among cell lines. For example, in pancreatic cancer cells, LDHB expression is significantly increased in PANC-1, MDA28, BxPC-3, Hs766T, but not expressed in CaPan-1, AsPC-1, MiaPaca-2, and PATU8902 pancreatic cancer cell lines[35]. Reports show that LDHB is highly expressed in basal-like breast cancer cell lines such as MDA-MB-231, MDA-MB-436, but not expressed in breast ductal carcinoma T-47D cell lines[36]; while for MCF-7 cell lines, reports on LDHB expression are inconsistent. Brown et al.[37] found that LDHB expression was absent, but Hussien et al.[38] confirmed that LDHB was predominantly expressed in MCF-7 cell lines. The variation in LDHB expression among different tumor cells (and even the absence of expression in some tumor cell lines) and the differences in specific energy metabolic modes of these cells are not well understood. One possible reason is that different environments/culture conditions lead to varying glycolytic dependencies among different tumor cells, and tumor cells with higher glycolytic dependencies also exhibit higher LDHB expression.
The diversity of LDHB expression in different tumor cells determines its role varies among different tumors. Liao et al.[39] found that after studying 269 cases of clinical primary localized bladder urothelial carcinoma patients and five bladder urothelial carcinoma cell lines with different invasion capabilities, LDHB expression levels were inversely correlated with tumor malignancy, patient survival, and corresponding tumor cell invasion capabilities. However, Shi et al.[40] showed that higher LDHB activity correlates with faster proliferation of colorectal cancer.
3
Development of Antitumor Drugs Targeting LDH
Due to the special position of LDH in tumor cell energy metabolism, the development of antitumor drugs targeting LDH has attracted increasing interest from scientists. Due to the diversity of LDHB expression in different tumor cells, there are currently few selective inhibitors developed for LDHB. In contrast, LDHA is often highly expressed in tumors, and inhibiting LDHA will reduce the conversion of pyruvate to lactate within tumor cells, thereby cutting off the ATP supply to tumor cells, while normal cells can directly utilize pyruvate to synthesize ATP in mitochondria, so they may be less affected. In addition, clinical reports have shown that long-term abnormal LDHA activity (such as genetic deficiency of LDHA) only causes non-lethal pustular psoriasis[41], indicating that targeting LDHA as a new target for tumor prevention and treatment is not only highly feasible but may also have fewer adverse reactions.
Oxamate is a classic LDHA inhibitor, with its structure shown in Figure 2A. It exists in normal human tissues and can competitively inhibit LDHA activity by competing with its substrate pyruvate. Research results confirm that oxamate can inhibit the proliferation, invasion, and migration of various tumor cells, including breast cancer, liver cancer, and non-small cell lung cancer, and can effectively enhance the sensitivity of other antitumor drugs and radiotherapy[42], but oxamate’s inhibitory effect on LDHA is non-specific and relatively weak, requiring millimolar concentrations for in vitro antitumor effects, limiting its clinical application value. It is primarily used as a tool drug for LDHA.
| |
| Figure 2Chemical Structures of Several Common LDHA Inhibitors |
Gossypol is a natural polyphenolic aldehyde extracted from cotton seeds, with its structure shown in Figure 2B. It is well-known as a male contraceptive due to its extreme sensitivity to the spermatogenic epithelial cells in the testis. Gossypol is also a classic LDHA inhibitor that can inhibit the proliferation of various tumor cells and promote their apoptosis, exhibiting significant antitumor activity. Its inhibitory effect on LDHA is also non-specific, and the numerous adverse reactions of gossypol limit its clinical application. An early phase I/II clinical trial reported[42] that the maximum tolerated dose of oral gossypol is 40 mg/d, with common adverse reactions including fatigue, nausea, vomiting, diarrhea, taste abnormalities, etc. Increasing the dose can also lead to more severe adverse reactions such as arrhythmias, hypokalemia, renal failure, muscle weakness, and even muscle paralysis.
FX11 is another recognized strong LDHA inhibitor derived from the structure of gossypol, with its structure shown in Figure 2C. Its IC50 against LDHA is about 50 nmol, and it can significantly inhibit the synthesis of lactate and ATP in tumor cells. It has been confirmed to inhibit tumor growth both in vitro and in vivo, but the catechol part of its structure limits its clinical application.
Additionally, some flavonoid compounds with polyphenolic structures, such as morin (structure shown in Figure 2D); epigallocatechin (structure shown in Figure 2E); also show certain LDHA inhibitory activity. In particular, galloflavin (structure shown in Figure 2F) can not only non-selectively inhibit LDHA but has also been confirmed to have good antitumor activity with relatively weak effects on normal cells and low toxicity[43].
4
Conclusion
Given the important role of LDH in tumor energy metabolism, especially the different energy metabolic modes between tumor cells and normal cells, it has become another potential target for tumor prevention and treatment. Over the past decade, a large number of LDHA inhibitors have been developed, and there is hope that by inhibiting LDHA, the ATP supply to tumor cells can be cut off, achieving antitumor effects. Although many of the above LDHA inhibitors have shown good antitumor effects at the cellular level, so far, no effective LDHA inhibitors have been used clinically. The possible reason may be related to the low bioavailability of the developed drugs. Zhang et al.[44] believe that due to the presence of a large number of cationic amino acid residues in the structure of LDHA, it is required that enzyme inhibitors ideally possess one or more negatively charged structures, such as -COO–, but inhibitors with such structures usually have poor permeability across cellular barriers.
Furthermore, Ždralević et al.[45] argue that only double genetic knockout of LDHA/B can completely suppress the “Warburg effect,” but even so, under aerobic conditions, some tumor cells can switch to OXPHOS to evade the drug’s effects and achieve rapid proliferation. Therefore, combining LDH inhibitors with inhibitors of other key enzymes in the glycolytic pathway, such as glucose transporter inhibitors and hexokinase inhibitors, may yield better antitumor effects. Thus, further in-depth research on the development of LDH inhibitors is necessary, and there may still be a considerable distance to go before they truly reach clinical application.
References
(Slide your finger in the box to browse)
[1] Warburg O. On the origin of cancer cells[J]. Science, 1956, 123(3191): 309-314.
[2] Felmlee MA, Jones RS, Rodriguez-Cruz V, et al. Monocarboxylate Transporters (SLC16): Function, Regulation, and Role in Health and Disease[J]. Pharmacol Rev, 2020, 72(2): 466-485.
[3] Pucino V, Cucchi D, Mauro C. Lactate transporters as therapeutic targets in cancer and inflammatory diseases[J]. Expert Opin Ther Targets, 2018, 22(9): 735-743.
[4] Contreras-Baeza Y, Sandoval PY, Alarcón R. Monocarboxylate transporter 4 (MCT4) is a high affinity transporter capable of exporting lactate in high-lactate microenvironments[J]. J Biol Chem, 2019, 294(52): 20135-20147.
[5] Guan X, Bryniarski MA, Morris ME. In Vitro and In Vivo Efficacy of the Monocarboxylate Transporter 1 Inhibitor AR-C155858 in the Murine 4T1 Breast Cancer Tumor Model[J]. AAPS J, 2018, 21(1): 3.
[6] Benjamin D, Robay D, Hindupur SK, et al. Dual Inhibition of the Lactate Transporters MCT1 and MCT4 Is Synthetic Lethal with Metformin due to NAD+ Depletion in Cancer Cells[J]. Cell Rep, 2018, 25(11): 3047-3058. e4.
[7] Srinivas SR, Gopal E, Zhuang L, et al. Cloning and functional identification of slc5a12 as a sodium-coupled low-affinity transporter for monocarboxylates (SMCT2)[J]. Biochem J, 2005, 392(Pt 3): 655-664.
[8] Hirschhaeuser F, Sattler UG, Mueller-Klieser W. Lactate: a metabolic key player in cancer[J]. Cancer Res, 2011, 71(22): 6921-6925.
[9] Haas R, Smith J, Rocher-Ros V, et al. Lactate regulates metabolic and pro-inflammatory circuits in control of T cell migration and effector functions[J]. PLoS Biol, 2015, 13(7): e1002202.
[10]Guan X, Morris ME. Pharmacokinetics of the Monocarboxylate Transporter 1 Inhibitor AZD3965 in Mice: Potential Enterohepatic Circulation and Target-Mediated Disposition[J]. Pharm Res, 2019, 37(1): 5.
[11]Hou L, Zhao Y, Song GQ, et al. Interfering cellular lactate homeostasis overcomes Taxol resistance of breast cancer cells through the microRNA-124-mediated lactate transporter (MCT1) inhibition[J]. Cancer Cell Int, 2019, 19: 193.
[12]Chen C, Zhu YH, Huang JA. Clinical evaluation of potential usefulness of serum lactate dehydrogenase level in follow-up of small cell lung cancer[J]. J Cancer Res Ther, 2018, 14(Supplement): S336-S340.
[13]Zhang X, Guo M, Fan J, et al. Prognostic significance of serum LDH in small cell lung cancer: A systematic review with meta-analysis[J]. Cancer Biomark, 2016, 16(3): 415-423.
[14] Xu XD, Shao SX, Jiang HP, et al. Warburg effect or reverse Warburg effect? A review of cancer metabolism[J]. Oncol Res Treat, 2015, 38(3): 117-122.
[15]Zong WX, Rabinowitz JD, White E. Mitochondria and Cancer[J]. Mol Cell, 2016, 61(5): 667-676.
[16]Epstein T, Gatenby RA, Brown JS. The Warburg effect as an adaptation of cancer cells to rapid fluctuations in energy demand[J]. PLoS One, 2017, 12(9): e0185085.
[17]de la Cruz-López KG, Castro-Muñoz LJ, Reyes-Hernández DO, et al. Lactate in the Regulation of Tumor Microenvironment and Therapeutic Approaches[J]. Front Oncol, 2019, 9: 1143.
[18]Brooks GA. The Science and Translation of Lactate Shuttle Theory[J]. Cell Metab, 2018, 27(4): 757-785.
[19]Liberti MV, Locasale JW. The Warburg Effect: How Does it Benefit Cancer Cells?[J]. Trends Biochem Sci, 2016, 41(3): 211-218.
[20]Cui J, Shi M, Xie D, et al. FOXM1 promotes the warburg effect and pancreatic cancer progression via transactivation of LDHA expression[J]. Clin Cancer Res, 2014, 20(10): 2595-2606.
[21]Jiang W, Zhou F, Li N, et al. FOXM1-LDHA signaling promoted gastric cancer glycolytic phenotype and progression[J]. Int J Clin Exp Pathol, 2015, 6: 6756-6763.
[22]Su Y, Yu QH, Wang XY, et al. JMJD2A promotes the Warburg effect and nasopharyngeal carcinoma progression by transactivating LDHA expression[J]. BMC Cancer, 2017, 17(1): 477.
[23]Jiang F, Ma S, Xue Y, et al. LDH-A promotes malignant progression via activation of epithelial-to-mesenchymal transition and conferring stemness in muscle-invasive bladder cancer[J]. Biochem Biophys Res Commun, 2016, 469(4): 985-992.
[24]Jiang P, Du W, Wu M. Regulation of the pentose phosphate pathway in cancer[J]. Protein Cell, 2014, 5(8): 592-602.
[25]Martinez-Outschoorn UE, Lin Z, Trimmer C, et al. Cancer cells metabolically’ fertilize’ the tumor microenvironment with hydrogen peroxide, driving the Warburg effect: Implications for PET imaging of human tumors[J]. Cell Cycle, 2011, 10(15): 2504-2520.
[26]Wang SJ, Xie J, Li JJ, et al. Cisplatin suppresses the growth and proliferation of breast and cervical cancer cell lines by inhibiting integrin β5-mediated glycolysis[J]. Am J Cancer Res, 2016, 6(5): 1108-1117.
[27]McCleland ML, Adler AS, Deming L, et al. Lactate dehydrogenase B is required for the growth of KRAS-dependent lung adenocarcinomas[J]. Clin Cancer Res, 2013, 19(4): 773-784.
[28]Li C, Chen Y, Bai PP, et al. LDHB may be a significant predictor of poor prognosis in osteosarcoma[J]. Am J Transl Res, 2016, 8(11): 4831-4843.
[29]Sun W, Zhang X, Ding X, et al. Lactate dehydrogenase B is associated with the response to neoadjuvant chemotherapy in oral squamous cell carcinoma[J]. PLoS One, 2015, 10(5): e0125976.
[30]Kumar S, Xie H, Scicluna P, et al. MiR-375 Regulation of LDHB Plays Distinct Roles in Polyomavirus-Positive and -Negative Merkel Cell Carcinoma[J]. Cancers (Basel), 2018, 10(11): 443.
[31]Kim JH, Kim EL, Lee YK, et al. Decreased lactate dehydrogenase B expression enhances claudin 1-mediated hepatoma cell invasiveness via mitochondrial defects[J]. Exp Cell Res, 2011, 317(8): 1108-1118.
[32]Wu GY, Yuan SC, Chen ZP, et al. The KLF14 Transcription Factor Regulates Glycolysis by Downregulating LDHB in Colorectal Cancer[J]. Int J Biol Sci, 2019, 15(3): 628-635.
[33]Maekawa M, Taniguchi T, Ishikawa J, et al. Promoter hypermethylation in cancer silences LDHB, eliminating lactate dehydrogenase isoenzymes 1-4[J]. Clin Chem, 2003, 49(9): 1518-1520.
[34]Leiblich A, Cross SS, Catto JW, et al. Lactate dehydrogenase-B is silenced by promoter hypermethylation in human prostate cancer[J]. Oncogene, 2006, 25(20): 2953-2960.
[35]Cui JJ, Quan M, Jiang WH, et al. Suppressed expression of LDHB promotes pancreatic cancer progression via inducing glycolytic phenotype[J]. Med Oncol, 2015, 32(5): 143.
[36] McCleland ML, Adler AS, Shang Y, et al. An integrated genomic screen identifies LDHB as an essential gene for triple-negative breast cancer[J]. Cancer Res, 2012, 72(22): 5812-5823.
[37]Brown NJ, Higham SE, Perunovic B, et al. Lactate dehydrogenase-B is silenced by promoter methylation in a highfrequency of human breast cancers[J]. PLoS One, 2013, 8(2): e57697.
[38]Hussien R, Brooks GA. Mitochondrial and plasma membrane lactate transporter and lactate dehydrogenase isoform expression in breast cancer cell lines[J]. Physiol Genomics, 2011, 43(5): 255-264.
[39]Liao AC, Li CF, Shen KH, et al. Loss of lactate dehydrogenase B subunit expression is correlated with tumour progression and independently predicts inferior disease-specific survival in urinary bladder urothelial carcinoma[J]. Pathology, 2011, 43(7): 707-712.
[40]Shi L, Yan H, An S, et al. SIRT5-mediated deacetylation of LDHB promotes autophagy and tumorigenesis in colorectal cancer[J]. Mol Oncol, 2019, 13(2): 358-375.
[41]Takeo N, Fujiwara S, Sakai T, et al. Hereditary Lactate Dehydrogenase M-subunit Deficiency With Late-Developing Pustular Psoriasis-Like Lesions[J]. J Dermatol, 2016, 43(12): 1429-1432.
[42]Koukourakis M, Tsolou A, Pouliliou S, et al. Blocking LDHA Glycolytic Pathway Sensitizes Glioblastoma Cells to Radiation and Temozolomide[J]. Biochem Biophys Res Commun, 2017, 491(4): 932-938.
[43]Van Poznak C, Seidman AD, Reidenberg MM, et al. Oral gossypol in the treatment of patients with refractory metastatic breast cancer: a phase Ⅰ/Ⅱ clinical trial[J]. Breast Cancer Res Treat, 2001, 66(3): 239-248.
[44]Zhang SL, He Y, Tam KY. Targeting cancer metabolism to develop human lactate dehydrogenase (hLDH) 5 inhibitors[J]. Drug Discov Today, 2018, 23(7): 1407-1415.
[45]Ždralević M, Brand A, Di Ianni L, et al. Double genetic disruption of lactate dehydrogenases A and B is required to ablate the “Warburg effect” restricting tumor growth to oxidative metabolism[J]. J Biol Chem, 2018, 293(41): 15947-15961.
(This article is an original article published in the Journal of Cancer Prevention and Treatment; please indicate the author and source when reprinting.)
