 A Bioinspired Lipopeptide Surmounts Therapeutic Dilemmas of Gram-Positive and Gram-Negative Polymicrobial InfectionJournal of the American Chemical Society ( IF 15.6 ) Pub Date : 2025-09-23DOI: 10.1021/jacs.5c10219
A Bioinspired Lipopeptide Surmounts Therapeutic Dilemmas of Gram-Positive and Gram-Negative Polymicrobial InfectionJournal of the American Chemical Society ( IF 15.6 ) Pub Date : 2025-09-23DOI: 10.1021/jacs.5c10219
Due to the lack of effective antimicrobial agents, especially those targeting Gram-positive and Gram-negative bacteria, polymicrobial infections pose significant therapeutic challenges. It is hypothesized that reasonably designed antimicrobial agents targeting both the outer and inner membranes of these two structurally distinct bacteria could provide solutions to the therapeutic dilemmas of polymicrobial infections.
Through lipid screening, a promising lipid scaffold was identified that can disrupt the membrane systems of both Gram-positive and Gram-negative bacteria. This paper developed a chemical optimization strategy to design a conical lipopeptide LP3K with broad-spectrum antibacterial activity, effective against polymicrobial infections without inducing resistance. We further elucidated the structure-activity relationship and antibacterial mechanism of LP3K. In a mouse model of polymicrobial infection, LP3K exhibited significant antibacterial and therapeutic effects, with confirmed in vivo biosafety. Our findings highlight the potential of lipopeptides as clinically translatable antibacterial agents for the effective treatment of polymicrobial infections.
In recent decades, bacterial infections have become a severe global health issue and are one of the leading causes of death worldwide. (1,2) Despite significant progress in the development of novel antimicrobial agents, (3−12) polymicrobial infections (especially those involving both Gram-positive and Gram-negative bacteria) still present enormous therapeutic challenges. (13,14) The lack of effective therapies for polymicrobial infections not only exacerbates the severity of diseases but also accelerates the emergence of antimicrobial resistance. (15−19) The main obstacle lies in the significant differences in the antibacterial efficacy of antibiotics against Gram-positive and Gram-negative bacteria, even for established broad-spectrum antibiotics. (20,21)
The structural differences between Gram-positive and Gram-negative bacteria pose a severe therapeutic dilemma. (21,22) Gram-positive bacteria have a thick peptidoglycan layer surrounding the inner membrane, while Gram-negative bacteria possess an additional outer membrane rich in lipopolysaccharides. (23 – 25) These structural differences lead to different therapeutic targets and bioavailability of antibiotics. (21) Notably, bacterial membranes are an attractive target for the development of novel antimicrobial agents that may exhibit rapid bactericidal action, multifaceted antibacterial mechanisms, and reduced potential for resistance development. (26 – 28) However, antimicrobial agents targeting membranes often exhibit different activities against Gram-positive and Gram-negative bacteria, complicating their application in polymicrobial infections. (26) For instance, polymyxin is very effective against Gram-negative bacteria by targeting the outer membrane but has limited efficacy against Gram-positive bacteria. (29) Furthermore, the structure-activity relationships of membrane-active compounds against these different bacterial types remain unclear, emphasizing the need for further research to optimize treatment strategies for polymicrobial infections. (26,30,31)
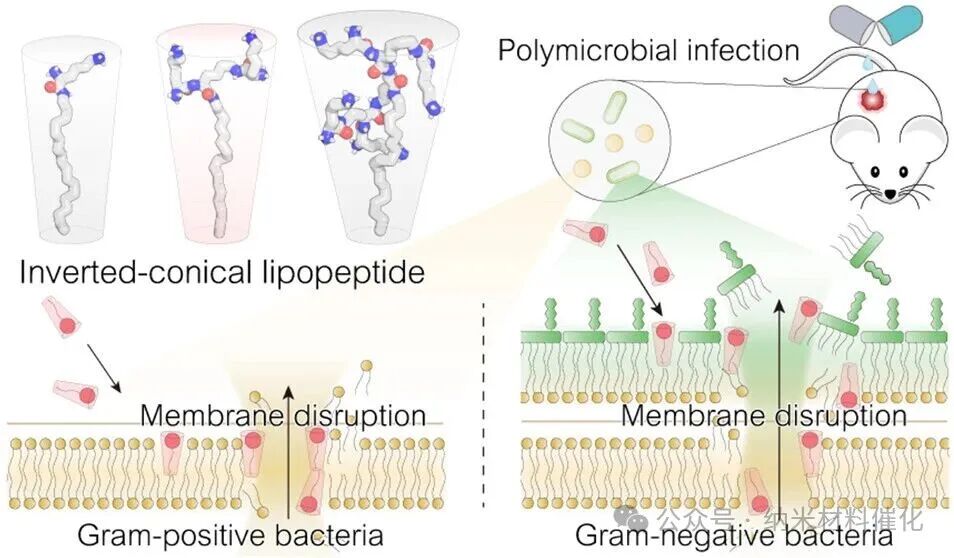
Bacterial membranes are primarily composed of cone-shaped lipids with large hydrophobic tails, such as lipopolysaccharides, phosphatidylethanolamine, and cardiolipin. (32,33) In nature, the differences in lipid types are crucial for membrane deformation processes (including pore formation, fusion, and fission). (34,35) Interestingly, the increased proportion of inverted cone-shaped lipids in lipid bilayers is associated with enhanced membrane permeability due to geometric antagonism. (35-37) Inspired by these natural phenomena, this study aims to elucidate the structure-activity relationships that are critical for the rational design of bioinspired antibacterial agents targeting polymicrobial infections (Figure 1A). This strategy includes screening common lipids to identify those with strong membrane-disrupting activity against both Gram-positive and Gram-negative bacteria, utilizing natural amino acids to fine-tune lipid shapes, revealing potential antibacterial mechanisms, and addressing the urgent therapeutic challenges posed by polymicrobial infections.
Experimental Results
Rational Design of Antibacterial Lipopeptides
The targeted action against bacterial cell membranes holds promise for the development of broad-spectrum, low-resistance, and bactericidal drugs, but overcoming the structural differences between Gram-positive and Gram-negative bacterial cell membranes remains challenging. By screening common compounds, lipids with strong interactions with bacterial cell membranes were identified (Figure S1). Given that Staphylococcus aureus (S. aureus ATCC29213) and Pseudomonas aeruginosa (P. aeruginosa ATCC27853) have typical cell membrane structures and often co-occur in clinical mixed infections, these two were selected as model strains. (38,39)
It was found that oleamide (OA) uniquely disrupts the cell membranes of both S. aureus and P. aeruginosa at concentrations above 16 μM, as assessed using propidium iodide (PI) as a membrane integrity probe, exhibiting concentration and time-dependent characteristics (Figure 1B and S2–S4). Meanwhile, octylamine and hexadecylamine showed differential membrane-disrupting activity in Gram-positive and Gram-negative bacteria, while other lipids did not exhibit significant membrane-disrupting effects. The membrane-disrupting ability of oleamide (OA) may be attributed to its lipid chain length, cis-unsaturated structure similar to bacterial lipids, and its positively charged hydrophilic head group. (40) However, the minimum inhibitory concentration (MIC) of OA against S. aureus is 128 μM, while the inhibitory concentration against P. aeruginosa exceeds 256 μM (Figure S5). Time-killing assays indicate that 256 μM of OA has limited bactericidal activity against both S. aureus and P. aeruginosa (Figure S6). Additionally, negatively charged lipids with the same hydrocarbon chain length as OA, such as oleic acid, exhibit very weak membrane-disrupting abilities against both Gram-positive and Gram-negative bacteria. The insufficient antibacterial activity and poor solubility of oleamide hinder its therapeutic potential, necessitating rational optimization.
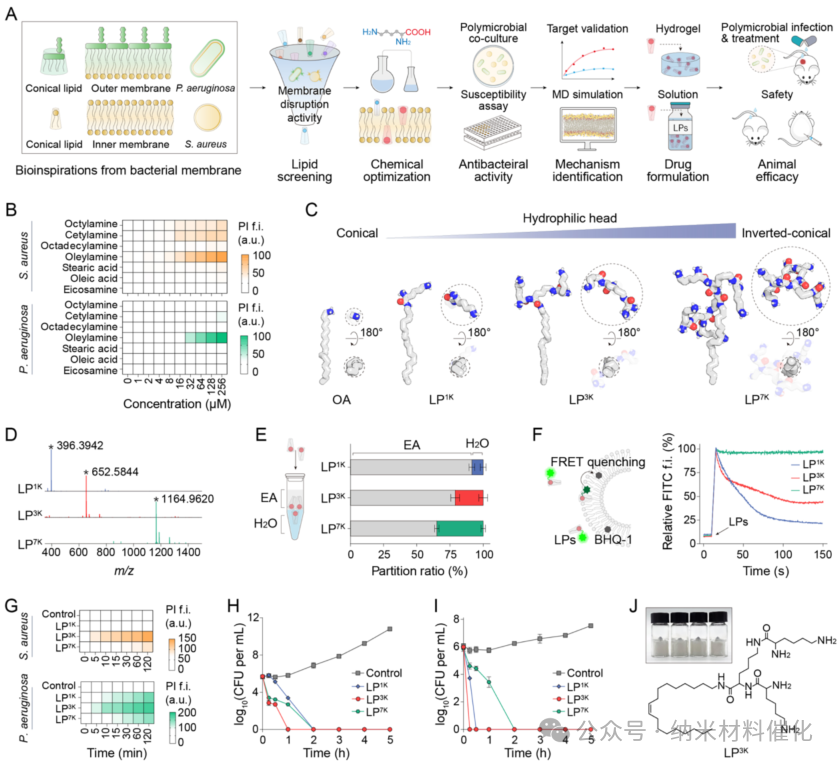
Figure 1. Development of Antibacterial Lipopeptides Based on Biological Principles. (A) Rational design of inverted cone-shaped lipopeptides targeting the cell membranes of Gram-positive and Gram-negative bacteria. (B) Membrane-disrupting activity of lipopeptides against S. aureus and P. aeruginosa at different concentrations over 1 hour, assessed using the PI probe. For example, fluorescence intensity. (C) Molecular dynamics simulations of OA, LP1K, LP3K, and LP7K in a simulated physiological environment after free evolution. (D) Mass spectra of lipopeptides LP1K (calculated [M + H]+ mass: 396.3948), LP3K (calculated [M + H]+ mass: 652.5848), and LP7K (calculated [M + H]+ mass: 1164.9646). Peaks marked with asterisks represent the corresponding [M + H]+ ions of each lipopeptide. (E) Distribution behavior of lipopeptides between ethyl acetate (EA) and water (H2O). (F) Membrane insertion ability of lipopeptides analyzed through FRET quenching. Arrows indicate the point of lipopeptide addition. (G) Membrane-disrupting effects of lipopeptides in Gram-positive and Gram-negative bacteria, assessed using the PI probe. Time-killing activity of lipopeptides under 2× MIC conditions against (H) S. aureus and (I) P. aeruginosa. (J) Chemical structure diagram of LP3K and a gram-scale photograph. Data are presented as mean ± standard deviation, with a sample size of 3.
Inspired by the significant interaction between oleamide and bacterial membranes, rational design of antibacterial agents was conducted to enhance their membrane-disrupting ability, antibacterial activity, and druggability. Given that negatively charged cone-shaped lipids are major components of bacterial cell membranes, the amine of OA was deliberately used as a branching center, and lysine (K), a basic amino acid, was used as a building block to synthesize inverted cone-shaped lipopeptides (LP, Figure 1C). (41 – 43) This unique branched peptide structure allows for rapid expansion of the hydrophilic head, introducing a large number of positively charged terminal amines from lysine residues, and exhibits higher molecular stability compared to traditional peptide antibiotics. (32,44,45) All-atom molecular dynamics (MD) simulations show that OA adopts a cone-shaped structure, while the first-generation lipopeptide (LP1K) transitions to an inverted cone-shaped structure. With the increase in the generation of branched peptides, the hydrophilic head gradually expands, with the second-generation lipopeptide (LP3K) and third-generation lipopeptide (LP7K) exhibiting increasingly pronounced inverted cone-shaped structures. The ratio of soluble surface area between the hydrophilic head and hydrophobic tail shows a consistent upward trend from LP1K to LP7K (Figure S7).
Next, these lipopeptide compounds were synthesized, and their detailed chemical structures and synthesis routes are provided in the supplementary information. The molecular structures were confirmed through 1H NMR spectroscopy and mass spectrometry, yielding precise mass-to-charge ratios of 396.3942 for LP1K, 652.5844 for LP3K, and 1164.9620 for LP7K (Figure 1D and S8 – S13). Additionally, high-performance liquid chromatography confirmed the high purity of all synthesized lipopeptides, exceeding 99%. The partition coefficients of LP1K, LP3K, and LP7K between the organic phase (ethyl acetate) and aqueous phase (H2O) were 10.72, 3.74, and 1.86, respectively (Figure 1E). These results indicate that the polarity significantly increases from LP1K to LP7K, and the morphology transitions to inverted cone-shaped. The calculated logP values of the lipopeptides confirm the increase in polarity from LP1K to LP7K (6.16, 4.12, and 0.04), attributed to the differences in branched peptides. The critical micelle concentration (CMC) also indicates that as the hydrophilic peptide head transitions from LP1K to LP7K, the hydrophilicity of the LP increases, while its aggregation ability in aqueous solution decreases (Figure S14).
A simulated bacterial membrane composed of phosphatidylglycerol (POPG) and phosphatidylcholine (POPC) was constructed, encapsulating a black hole quencher 1 (BHQ-1) within the bilayer structure. When fluorescently labeled lipopeptides (LPs) were inserted into the membrane, the fluorescence signal was quenched to BHQ-1 through fluorescence resonance energy transfer (FRET), allowing for the assessment of membrane insertion ability (Figure 1F). LP1K and LP3K triggered rapid fluorescence quenching, indicating strong membrane insertion ability. In contrast, LP7K exhibited poor insertion ability in artificial membranes, likely due to its excessively large hydrophilic head. These findings confirm that an appropriate proportion of inverted cone shape enhances the ability of lipopeptides to disrupt membranes, while the excessively large hydrophilic head of LP7K weakens this effect. This conclusion aligns with the superior membrane-disrupting ability exhibited by LP3K in both Gram-positive and Gram-negative bacteria, as evidenced by the PI membrane permeability phenomenon (Figure 1G). The MIC values of LP1K, LP3K, and LP7K indicate that they exhibit significant inhibitory effects against both S. aureus and P. aeruginosa, with effects far exceeding those of oleamide after drug optimization (Figure S15). Furthermore, colony-forming unit (CFU) counts revealed that LP3K has a significant bactericidal advantage against both Gram-positive and Gram-negative bacteria, capable of eliminating S. aureus within 60 minutes and P. aeruginosa within 15 minutes (Figure 1H, I). LP1K and LP7K also demonstrated considerable bactericidal capabilities, surpassing many widely used antibiotics such as vancomycin, polymyxin B, amoxicillin, and azithromycin (Figure S16). Additionally, we achieved large-scale synthesis of these lipopeptides with high yields. Overall, the large-scale synthesis, excellent membrane-disrupting ability, and impressive bactericidal performance make LP3K a promising candidate for a new generation of antibacterial drugs (Figure 1J).
Subsequent analyses indicated that LP3K exhibits broad-spectrum antibacterial activity, with essentially equivalent antibacterial effects against both Gram-positive and Gram-negative pathogens (Table 1). This sharply contrasts with the pronounced selectivity exhibited by vancomycin and polymyxin B. Moreover, commonly used broad-spectrum antibiotics such as amoxicillin, tetracycline, ciprofloxacin, and gentamicin also show significant differences in MIC values between Gram-positive and Gram-negative bacteria (Figure S17). Notably, LP3K also exhibits inhibitory effects against methicillin-resistant Staphylococcus aureus (MRSA) and clinical isolates. Remarkably, all MIC values of LP3K against both Gram-positive and Gram-negative bacteria are within the same order of magnitude, which is significantly different from traditional antibiotics. These findings strongly suggest that LP3K can overcome the therapeutic barriers posed by Gram-positive and Gram-negative bacteria in polymicrobial infections.
Antibacterial Lipopeptides Against Polymicrobial Co-cultures
Polymicrobial mixed infections present severe therapeutic challenges, as few antimicrobial agents can clear polymicrobial infections containing both Gram-positive and Gram-negative bacteria. To simulate the environment of polymicrobial mixed infections, S. aureus (ATCC29213) and P. aeruginosa (ATCC27853) were co-cultured at equivalent colony-forming units. The MIC of LP3K against the polymicrobial co-culture was determined to be 16 μM, significantly lower than the MIC values of vancomycin (>256 μM) and polymyxin B (128 μM) (Figure 2A). Notably, this MIC value is below the critical micelle concentration (CMC) of LP3K, indicating that the lipopeptide primarily exerts its antibacterial activity in a non-assembled monomeric form. Due to the inhibitory effect of LP3K, the turbidity of the polymicrobial co-culture treated with LP3K was comparable to that of the blank medium, while treatments with vancomycin or polymyxin B at the same time point showed significant turbidity, indicating substantial bacterial proliferation (Figure 2B).
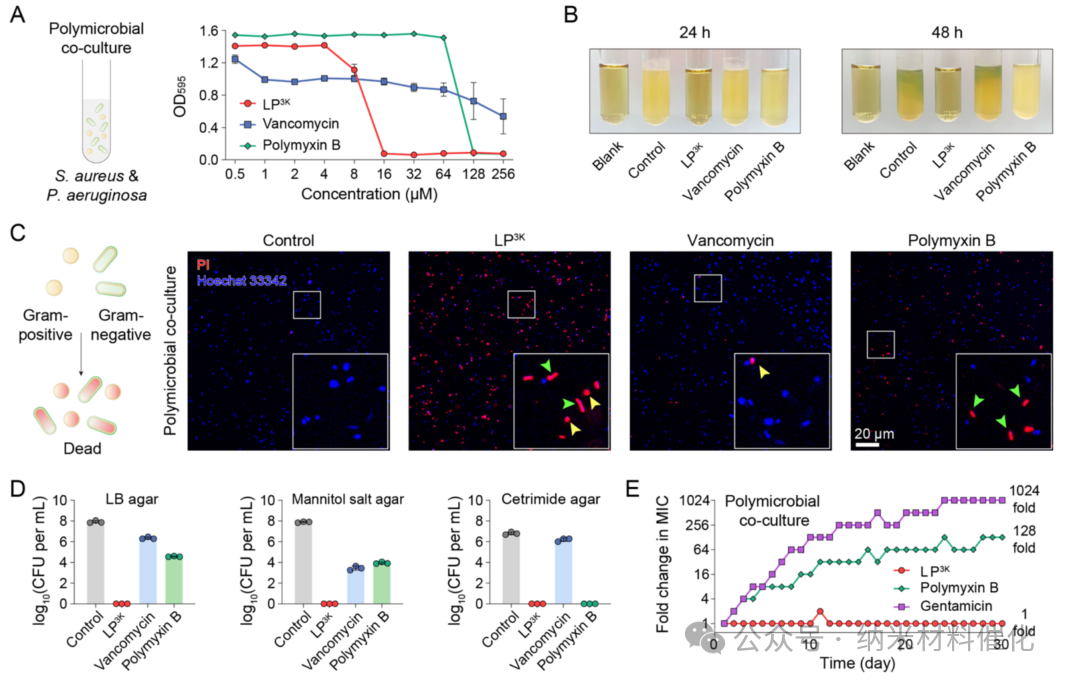
Figure 2. Antibacterial Effects of LP3K on Polymicrobial Co-cultures. (A) Inhibition of polymicrobial co-cultures of S. aureus and P. aeruginosa by LP3K, vancomycin, and polymyxin B over 24 hours at concentrations ranging from 0.5 to 256 μM. (B) Images of polymicrobial co-cultures after 24 and 48 hours of incubation in media containing 64 μM of the antibacterial agent. Bacterial culture media without any treatment (blank control) and untreated bacterial cultures (control group) were used as controls. (C) Confocal microscopy images of polymicrobial co-cultures stained with PI (red) and Hoechst 33342 (blue), as well as after exposure to the indicated antibacterial agents at 64 μM for 1 hour. (Scale bar: 20 μm) (D) Bacterial counts in polymicrobial co-cultures after exposure to the indicated antibacterial agents at 64 μM for 3 hours, determined by plating on LB agar, mannitol salt agar (selective for S. aureus), and ertapenem agar (selective for P. aeruginosa). (E) Development of resistance in polymicrobial co-cultures under sub-MIC levels of LP3K, polymyxin B, and gentamicin. For gentamicin, 1024 times the minimum inhibitory concentration was the highest concentration tested. Data are presented as mean ± standard deviation, n = 3.
Next, the dual staining method with PI and Hoechst 33342 confirmed that LP3K exhibits bactericidal activity: within just 1 hour, over half of the bacteria exhibited a PI-positive state, a result significantly superior to that of vancomycin and polymyxin B. Fluorescence spectral analysis further indicated that through confocal laser scanning microscopy (CLSM), LP3K could induce substantial cell death in both cocci and rod-shaped bacteria, while vancomycin failed to affect any rod-shaped cells, and polymyxin B was ineffective against cocci (Figure 2C and S18). Plating experiments conducted on LB agar plates quantified the bactericidal effects against the polymicrobial co-culture, revealing that LP3K completely eliminated the bacteria after 3 hours of incubation (Figure S19). As expected, neither vancomycin nor polymyxin B could eradicate this polymicrobial co-culture at the same concentration, with bacterial counts remaining at 2.33×10^6 and 3.81×10^4 CFU mL^-1, respectively (Figure 2D).
Additionally, selective mannitol salt agar demonstrated that LP3K effectively kills S. aureus, while both vancomycin and polymyxin B failed to eliminate S. aureus in this polymicrobial co-culture. Similarly, selective ciprofloxacin agar confirmed that LP3K completely cleared Gram-negative bacteria in the polymicrobial co-culture. Furthermore, time-killing assays indicated that LP3K thoroughly eliminated the polymicrobial culture within 30 minutes, while the bactericidal activity of vancomycin and polymyxin B was significantly weaker (Figure S20). Even the combination of vancomycin and polymyxin B required 4 hours to eradicate the bacterial population, indicating that their bactericidal efficiency is inferior to that of LP3K. Moreover, the addition of LP3K to various agar plates completely inhibited the proliferation and spread of the polymicrobial culture (Figure S21).
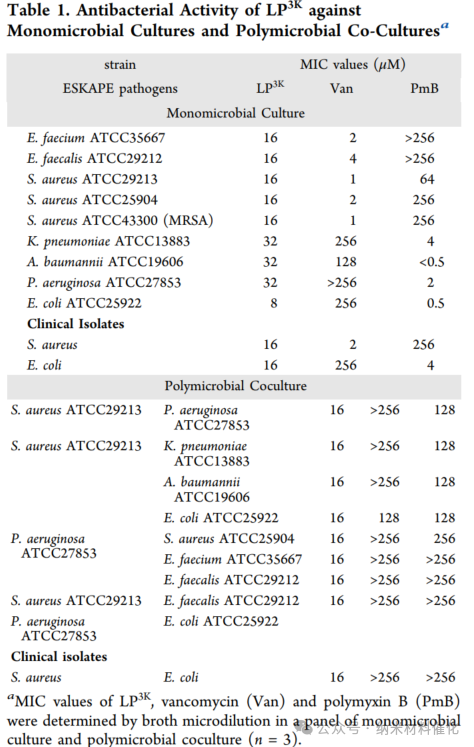
Furthermore, the minimum inhibitory concentration (MIC) of LP3K consistently remained at 16 μM in polymicrobial co-cultures of eight Gram-positive and Gram-negative bacteria (including ESKAPE pathogens and clinical isolates) (details in Table 1). This observation highlights the broad-spectrum activity of LP3K against polymicrobial infections, even when multiple strains are involved. In contrast, both vancomycin and polymyxin B struggled to suppress the growth of polymicrobials. More concerning is the significant development of resistance, as evidenced by the 128-fold increase in the MIC of polymyxin B within 30 days (Figure 2E). Encouragingly, LP3K exhibited almost no development of resistance during the same time period. In comparison, gentamicin, a frontline clinical antibiotic for treating polymicrobial infections, also saw its MIC increase by over 1024-fold within 30 days.
Overall, LP3K demonstrated significant efficacy against polymicrobial infections, underscoring the potential of a powerful antibacterial agent. Polymicrobial infections are often associated with biofilm formation, which confers considerable resistance to conventional antimicrobial therapies. To evaluate the antibiofilm potential of LP3K, its ability to prevent biofilm formation by polymicrobial cultures consisting of S. aureus and P. aeruginosa was first assessed. At concentrations as low as 16 μM, LP3K completely inhibited biofilm formation, outperforming vancomycin and polymyxin B (Figure S24). Additionally, LP3K was able to clear mature polymicrobial biofilms (Figure S25), demonstrating superior efficacy compared to clinically used antibiotics. Notably, LP3K exhibited robust bactericidal activity against Gram-positive and Gram-negative bacteria embedded within established biofilms (Figure S26). These findings suggest that LP3K holds great potential in combating polymicrobial infections associated with biofilm formation and disrupting established microbial communities.
Antibacterial Mechanism Against Polymicrobial Co-cultures
Significant interactions between LP3K and pathogenic bacteria were observed through microscale thermophoresis (MST), involving various strains of S. aureus, P. aeruginosa, and their mixed cultures. In contrast, vancomycin interacted only with the Gram-positive bacterium S. aureus, while polymyxin B interacted solely with the Gram-negative bacterium P. aeruginosa (Figure 3A and S27). The dissociation constants (Kd) indicated that LP3K exhibited comparable Kd values with each strain and their mixed bacteria, all within the same order of magnitude (Figure 3B). However, vancomycin and polymyxin B displayed significant differences in Kd values across different Gram-positive and Gram-negative bacteria, with Kd values for the polymicrobial co-culture being undetectable. Subsequently, flow cytometry elucidated the binding kinetics of the antibacterial agents with bacterial cells (Figure 3C). LP3K rapidly bound to the polymicrobial population, achieving a maximum binding rate of 92.63 ± 1.53% of bacteria at 180 minutes, indicating its rapid and strong binding. In contrast, the binding rates of vancomycin and polymyxin B to bacteria were slower and less effective.
Confocal laser scanning microscopy (CLSM) images revealed that LP3K exhibited strong interactions with both cocci and rod-shaped bacteria, while vancomycin failed to bind to rod-shaped bacteria, and polymyxin B showed weak binding to cocci (Figure 3D and S29). The zeta potential of bacterial cells shifted from negative to positive under the concentration-dependent action of LP3K, reflecting its interaction and binding with the negatively charged bacterial surface. These results indicate that LP3K can effectively bind to both Gram-positive and Gram-negative bacteria, providing a strong theoretical basis for its potent antibacterial effects against polymicrobial co-cultures.
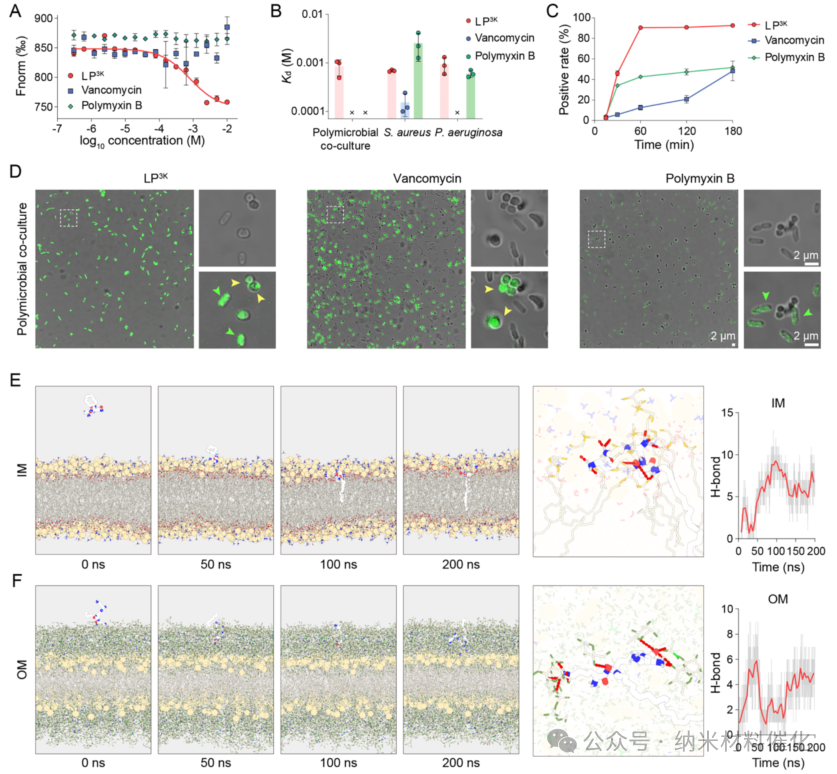
Figure 3. Binding of LP3K to Polymicrobial Co-cultures. (A) Quantitative analysis and (B) Kd values of LP3K, vancomycin, and polymyxin B binding affinity to mixed cultures of S. aureus and P. aeruginosa, determined by microscale thermophoresis. Kd values were fitted using MO affinity analysis (NanoTemper) software. (C) Binding rates of the mixed cultures assessed by flow cytometry in the presence of fluorescently labeled LP3K, vancomycin, and polymyxin B analogs. (D) Confocal laser scanning microscopy images of polymicrobial co-cultures containing the indicated antibacterial agents. Scale bar: 2 μm. Molecular dynamics simulations of LP3K binding to (E) the inner membrane (IM) and (F) the outer membrane (OM) were conducted after 200 nanoseconds of free evolution. The images show the distance of LP3K from the membrane and the formation of typical hydrogen bonds at 0, 50, 100, and 200 nanoseconds. The line graph describes the relationship between the number of hydrogen bonds and simulation time. Data are presented as mean ± standard deviation, n = 3.
Moreover, molecular dynamics simulations demonstrated that the inverted cone-shaped LP3K exhibits significant interactions with the model of the bacterial inner membrane, a feature that is prevalent in both Gram-positive and Gram-negative bacteria. The amphiphilic nature of LP3K allows it to interact with the inner membrane in approximately 7.6 seconds, leading to stable insertion into the lipid bilayer after about 78.0 seconds (Figure 3E and S31A). LP3K forms up to 13 hydrogen bonds with the inner membrane model, primarily contributed by the branched peptide. Similarly, LP3K also exhibited stable interactions with the outer membrane, characteristic of Gram-negative bacteria, reaching a maximum number of 9 hydrogen bonds after approximately 29.5 seconds (Figure 3F and S31B). These phenomena provide compelling evidence for the supramolecular interactions between inverted cone-shaped LP3K and bacterial membranes (including hydrophobic and hydrogen bonding interactions), elucidating its effective binding capabilities to both Gram-negative and Gram-positive bacteria.
This carefully designed LP3K can generate strong interactions with S. aureus, P. aeruginosa, and their mixed cultures, prompting us to further investigate its antibacterial mechanism of membrane damage (Figure 4A). (46,47) Initially, the fluorescent probe 1-phenylnaphthylamine (NPN) was used to assess the integrity of the outer membrane. Upon disruption of the outer membrane, NPN enters the hydrophobic layer of the inner membrane, leading to a significant increase in fluorescence (Figure 4B). After treatment with LP3K, the fluorescence intensity of NPN at 420 nm increased 2.3-fold in the polymicrobial co-culture, indicating substantial damage to the outer membrane.
Additionally, o-nitrophenyl-β-galactopyranoside (ONPG) was used to evaluate the integrity of the inner membrane. Notably, LP3K caused significant disruption of the inner membrane systems of S. aureus, P. aeruginosa, and the polymicrobial culture, as reflected by the increase in optical density (OD420) at 420 nm (Figure 4C). Real-time detection of PI influx demonstrated that LP3K rapidly and strongly compromised the integrity of bacterial cell membranes, including both inner and outer membranes, even in the context of polymicrobial co-cultures (Figure 4D).
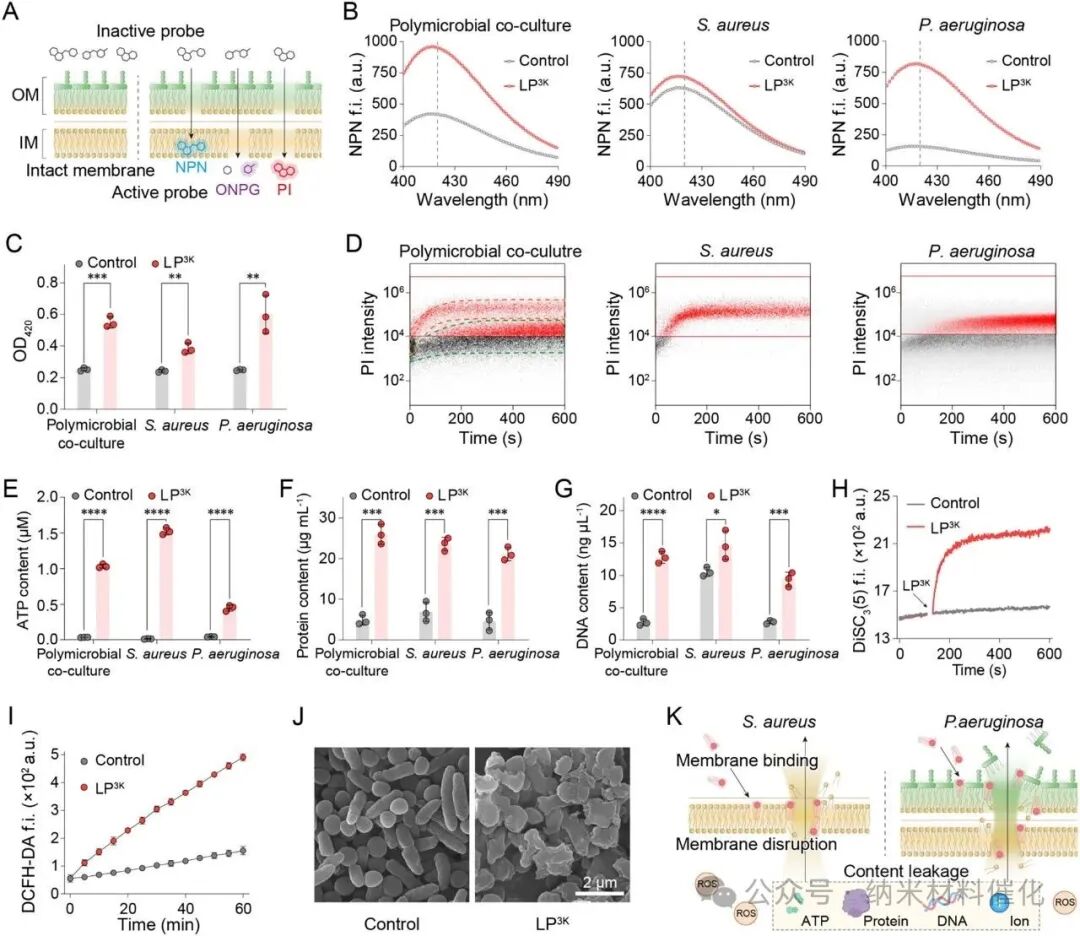
Figure 4. LP3K Disrupts Bacterial Outer and Inner Membranes. (A) Schematic diagram illustrating the study of bacterial inner and outer membrane integrity. (B) NPN fluorescence spectra showing the permeability of the outer membrane in mixed cultures of S. aureus and P. aeruginosa under the influence of 64 μM LP3K (excitation light at 350 nm). Differences in fluorescence intensity were analyzed at 420 nm. (C) OD420 indicating damage to the inner membrane in mixed cultures exposed to 64 μM LP3K (mediated by ONPG hydrolysis). (D) Real-time flow cytometry density plots showing PI influx, indicating damage to the bacterial membranes in the mixed cultures. Red and green dashed lines indicate the aggregation of the two bacterial species. (E)(F)(G) Leakage of ATP, proteins, and DNA from the supernatant of mixed cultures after exposure to 64 μM LP3K for 1 hour. (H) Membrane depolarization experiments of mixed cultures detected using the DiSC3(5) probe during treatment with 64 μM LP3K. Arrows indicate the addition of LP3K. (I) Measurement of oxidative stress levels in mixed cultures using the ROS probe DCFH-DA (excitation light at 488 nm, emission light at 525 nm) during treatment with 64 μM LP3K. (J) Representative scanning electron microscopy images of polymicrobial co-cultures after exposure to 64 μM LP3K for 1 hour. Scale bar: 2 μm. (K) Schematic diagram of the antibacterial mechanism of LP3K against polymicrobial co-cultures of Gram-positive and Gram-negative bacteria. Data are presented as mean ± standard deviation, n = 3. Statistical significance was determined by unpaired t-test, ****P < 0.0001; ***P < 0.001; **P < 0.01; *P < 0.05.
As shown in Figure 4E, LP3K treatment significantly increased the release of endogenous ATP in S. aureus, P. aeruginosa, and polymicrobial co-cultures, with ATP release being a key indicator of bactericidal effects. Furthermore, LP3K also promoted the efflux of endogenous proteins from S. aureus and P. aeruginosa, with higher levels of protein leakage observed in the polymicrobial co-culture (Figure 4F). Additionally, LP3K treatment led to a significant increase in bacterial DNA leakage across all groups, indicating extensive membrane damage that allowed macromolecules to pass through (Figure 4G). As expected, LP3K rapidly induced membrane depolarization in the polymicrobial co-culture, as evidenced by the rapid increase in fluorescence intensity of the lipophilic potential probe DiSC3(5) (Figure 4H).
LP3K-mediated membrane damage also resulted in oxidative stress across all treatment groups, as reflected by the intense fluorescence generated by the reactive oxygen species (ROS) probe 2′,7′-dichlorofluorescein diacetate (DCFH-DA) (Figure 4I and S35). Scanning electron microscopy (SEM) images revealed a loss of typical cocci and rod-shaped morphologies, indicating severe membrane damage and cell lysis following LP3K treatment (Figure 4J and S36).
In summary, these findings indicate that inverted cone-shaped lipopeptides effectively disrupt the outer and inner membranes of pathogenic bacteria, leading to the leakage of endogenous molecules and ultimately resulting in bacterial death (Figure 4K). These mechanisms successfully overcome the therapeutic barriers posed by Gram-positive and Gram-negative bacteria in polymicrobial co-cultures.
Lipopeptides for the Treatment of Polymicrobial Infections
To further explore the in vivo antibacterial effects of lipopeptides against polymicrobial infections, we established an animal model by inoculating equal amounts of 1×10^6 CFU of S. aureus and 1×10^6 CFU of P. aeruginosa on the skin of BALB/c mice. To enhance treatment compliance and translational potential, LP3K was loaded into a biocompatible hydrogel as a wound dressing formulation (Figure 5A-C and S44). (47,48) Four hours after polymicrobial infection, LP3K, vancomycin, and polymyxin B were administered at a dose of 20 mg per kg body weight (Figure 5D).
Similar purulent conditions were observed in the vancomycin and polymyxin B treatment groups, reflecting their inability to combat polymicrobial infections. In contrast, only a small amount of purulent fluid was observed in the mice treated with LP3K, indicating its significant bactericidal effects against polymicrobial infections. To quantify the therapeutic effects, full-thickness skin samples were collected from the wounds for plating. In the untreated group, the total bacterial load within the tissue was 8.33×10^8 CFU/g. However, even with treatment using vancomycin or polymyxin B, bacterial counts remained as high as 1.86×10^7 CFU/g and 4.16×10^7 CFU/g, respectively. Encouragingly, only 4.39×10^5 CFU/g of bacteria were detected at the wounds of mice with polymicrobial infections treated with LP3K. Compared to the groups treated with vancomycin and polymyxin B, LP3K demonstrated over 40-fold and 90-fold reductions in bacterial counts against polymicrobial infections, respectively. Selective culture media revealed that LP3K exhibited stronger antibacterial activity against Gram-positive bacteria than vancomycin, while its efficacy against Gram-negative bacteria in polymicrobial infections was comparable to that of polymyxin B (Figure 5E).
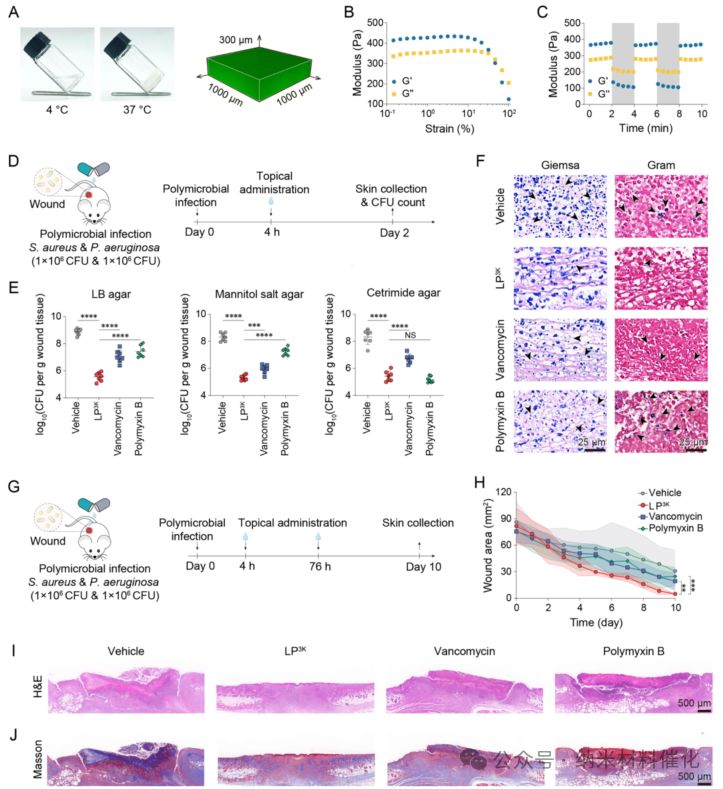
Figure 5. Antibacterial Effects of LP3K in Polymicrobial Infections. (A) Temperature sensitivity of hydrogel dressings loaded with LP3K and Z-stack scanning images of fluorescently labeled LP3K analogs loaded in hydrogel dressings. The dressing remains liquid at 4°C and transitions to solid at 37°C. The green signal confirms the dispersion of LP3K within the dressing. (B) Stretching scan of the dressing at 37°C under a shear frequency of 10 rad s-1. (C) Time scan of the dressing at 37°C transitioning from lower shear strain (1%) to higher shear strain (100%) at a shear frequency of 10 rad s-1. (D) Schematic diagram of the short-term in vivo study using a mouse model with polymicrobial infections. (E) Bacterial loads in polymicrobial infection wounds after treatment with vehicle (hydrogel), LP3K, vancomycin, or polymyxin B, all at a dose of 20 mg per kg body weight. Bacterial counts were determined on LB, mannitol salt, and ertapenem agar plates. Seven mice per group. (F) Representative Giemsa and Gram staining images of polymicrobial infection wounds after short-term treatment. Arrows indicate bacteria. Scale bar: 25 μm. (G) Schematic diagram of the long-term in vivo study using a mouse model of polymicrobial infections. (H) Wound healing progress of polymicrobial infection mice treated with different drugs (lipopeptide LP3K, vancomycin, or polymyxin B, all at a dose of 20 mg per kg body weight). Seven mice per group. Representative (I) H&E staining images and (J) Masson’s trichrome staining images showing the wound conditions of polymicrobial infection mice under long-term different treatments. Scale bar: 500 μm. Data are presented as mean ± standard deviation. Statistical significance was determined by unpaired t-test, ****P < 0.0001; ***P < 0.001; **P < 0.01; NS, no significant difference.
Longitudinal tissue section analysis was conducted to assess the status of polymicrobial infections, using Giemsa staining to detect total bacteria and Gram staining to detect Gram-positive bacteria (Figure 5F). Giemsa staining of bacterial spots revealed that vancomycin and polymyxin B exhibited moderate antibacterial effects, while LP3K demonstrated more efficient antibacterial activity, clearing fewer bacteria.
To evaluate the efficacy of LP3K against polymicrobial infections, a long-term wound healing observation was conducted over 10 days, with two doses of 20 mg per kg body weight administered (Figure 5G). The wound healing rates in the vancomycin and polymyxin B groups were 74.84 ± 9.54% and 66.84 ± 14.59%, both lower than the solvent control group (64.06 ± 22.86%, see Figure 5H and S46B). These groups also exhibited greater individual variability in wound healing. H&E staining of wound site sections showed that LP3K treatment promoted nearly complete wound healing, with intact skin tissue structure, while the solvent control group exhibited significant skin damage (Figure 5I). However, unhealed skin damage was still observed in the vancomycin and polymyxin B groups, further demonstrating the antibacterial efficacy of LP3K in combating polymicrobial infections and promoting wound healing. Masson’s trichrome staining further indicated a greater deposition of collagen in the wound tissue of the LP3K treatment group (Figure 5J). In contrast, wounds in other groups with polymicrobial infections showed minimal collagen deposition and poor healing outcomes.
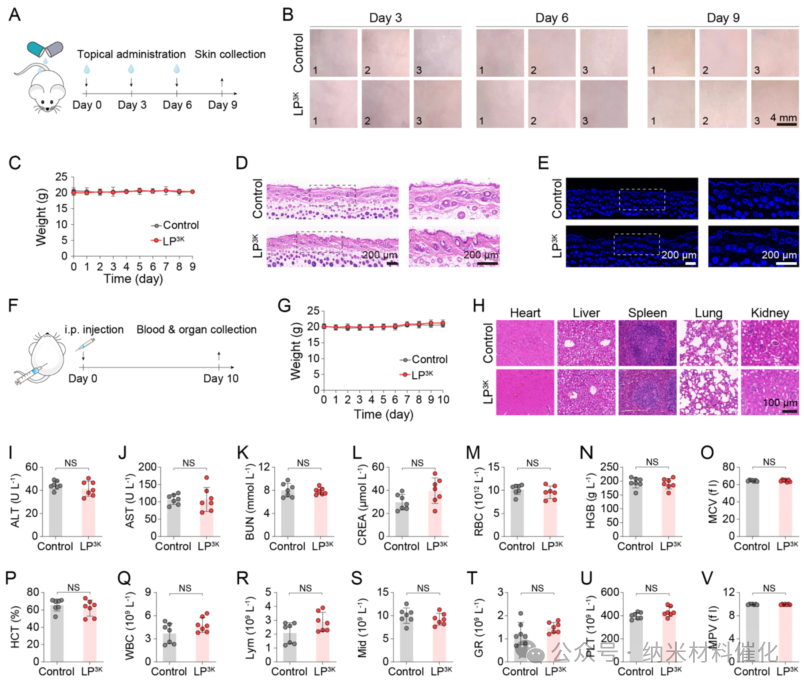
Figure 6. In Vivo Safety Study of LP3K. (A) Schematic diagram of the in vivo safety study of LP3K on mouse skin. (B) Representative images of the skin after each administration of LP3K. Physiological saline treatment served as a control. Scale bar: 4 mm. Images represent 3 mice per group. (C) Weight changes in mice during the skin safety study. Three mice were used per group. Representative (D) H&E staining images and (E) TUNEL staining images showing the skin of mice after three administrations of LP3K. Scale bar: 200 μm. (F) Schematic diagram of the in vivo safety study conducted via intraperitoneal injection (i.p.). Safety of LP3K administered intraperitoneally: (G) Weight changes, (H) Representative H&E staining images of major organs, (I) Plasma biochemical analysis of alanine aminotransferase (ALT), (J) Plasma biochemical analysis of aspartate aminotransferase (AST), (K) Plasma biochemical analysis of blood urea nitrogen (BUN) and (L) creatinine (CREA), as well as (M) red blood cells (RBC), (N) hemoglobin (HGB), (O) mean corpuscular volume (MCV), (P) hematocrit (HCT), (Q) white blood cells (WBC), (R) lymphocyte counts (Lym), (S) intermediate cell counts (Mid), (T) granulocyte counts (GR), (U) platelets (PLT), and (V) mean platelet volume (MPV). Scale bar: 100 μm. Seven mice per group. Data are presented as mean ± standard deviation. Statistical significance was determined by unpaired t-test, NS indicates no statistical significance.
Additionally, LP3K loaded in a hydrogel formulation was applied topically to the skin of BALB/c mice every 3 days for 9 days (Figure 6A). No signs of erythema, swelling, or inflammation were observed, indicating no significant local irritation (Figure 6B). No significant weight loss was observed throughout the experiment (Figure 6C). H&E staining of skin sections from LP3K-treated mice showed no significant differences in epidermal thickness and inflammatory cell infiltration compared to the control group (Figure 6D). No significant apoptosis was observed in the skin of TUNEL-stained samples (Figure 6E). Intraperitoneal administration of LP3K (20 mg per kg body weight, solution formulation) resulted in 100% survival and normal weight over 10 days (Figure 6F, G). H&E staining of major organs from LP3K-treated mice showed no signs of tissue damage (Figure 6H and S47). Furthermore, blood tests (including serum biochemical markers (ALT, AST, BUN, CREA, Figure 6I – L) and hematological parameters (Figure 6M – V and S48A – G)) revealed no significant differences between LP3K-treated mice and the saline-injected control group.
Conclusion
The increasing prevalence of polymicrobial infections, coupled with the severe shortage of effective antibacterial agents, underscores the urgent need for the development of effective antibacterial agents. The differences in cell membranes between Gram-positive and Gram-negative bacteria not only lead to differences in antibacterial effects but also represent key targets for effectively eliminating polymicrobial infections. Inspired by bacterial cell membranes, both Gram-positive and Gram-negative bacterial cell membranes are composed of cone-shaped lipids; moreover, the natural membrane deformation processes often contain more inverted cone-shaped lipids. By screening common lipids, oleamide was identified as a promising lipid scaffold due to its potential to disrupt the cell membranes of both Gram-positive and Gram-negative bacteria.
This paper developed a design strategy for inverted cone-shaped lipids, elucidating the structure-activity relationships that underpin the efficacy of lipopeptides. The optimization of this drug led to the discovery of lipopeptide candidates with powerful broad-spectrum antibacterial activity (capable of combating mixed bacterial populations) without the development of resistance being observed. Through molecular, cellular, and computational analyses, the bactericidal mechanisms of these lipopeptides were revealed: inverted cone-shaped LP3K effectively disrupts the outer and inner membranes of bacteria, leading to the leakage of important biomolecules and halting biosynthesis. In vivo, LP3K demonstrated superior efficacy in preventing bacterial colonization in polymicrobial infections compared to polymyxin B and vancomycin. Notably, no local or systemic toxicity was observed in mouse models.
Furthermore, these intriguing lipopeptides possess favorable physicochemical properties, and the production process and formulation development have been successfully achieved, further highlighting their druggability and translational potential, making them effective antibacterial agents against polymicrobial infections. Future research should focus on pharmacokinetics, pharmacodynamics, and more animal models of polymicrobial infections to facilitate the drug development process. A comprehensive exploration of the design and optimization of antibacterial lipopeptides can provide a novel strategy for combating complex infectious diseases, especially in the face of clinical challenges posed by multiple pathogens.
WeChat Official Account丨Nanomaterials Catalysis
Disclaimer: The copyright of this article belongs to the original author. Please contact for authorization if reprinting! Due to limited knowledge, there may be omissions and errors, and criticism and correction are welcome!
For collaboration inquiries, please contact us via private message.