Author Click HereLanFollow Us
Hello everyone, the article we are pushing today is titled “Structural Basis of High-Precision Protein Ligation and Its Application,” published in the Journal of the American Chemical Society in January 2025.
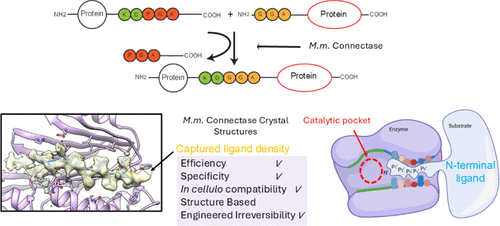
Enzyme-catalyzed protein modifications have become extremely valuable in various applications, outperforming chemical methods in terms of precision, coupling efficiency, and biocompatibility. Despite significant progress made by ligation enzymes (such as Sortase A and OaAEP1), their application in heterogeneous biological environments is still limited by the target sequence specificity. The Lupa’s team introduced a ligation enzyme (Connectase), a family of archaeal proteases modified for protein ligation, but its low sustained synthesis capability and lack of structural information hinder further engineering modifications for practical biological and biophysical applications. Here, we present the X-ray crystal structures of Mm ligase (MmCET from Methanococcus maripaludis) in both apo form and substrate-bound form. By comparing it with its inactive homolog MjCET (Methanococcus janaschii), we reveal the structural basis for the high-precision ligation activity of MmCET. The authors propose modifications to the N-terminal substrate recognition motif to inhibit the reversible protease activity of MmCET, thereby achieving high-precision protein ligation in complex biological environments (such as serum-containing cell cultures). To further demonstrate its enhanced sustained synthesis capability and precision, single-molecule protein unfolding experiments show that the optimized ligase used in conjunction with OaAEP1 (C247A) can perform stepwise tandem ligation of proteins, forming structurally defined protein polymers.
1
Screening of ATA
Understanding the structural features that distinguish ligation enzymes from proteases is key to advancing enzyme-catalyzed protein ligation technology. The study first elucidated the crystal structures of MmCET in its apo form (PDB code: 8JTU) and in the peptide substrate-bound form, focusing on the structural differences compared to its inactive homolog MjCET (Methanococcus janaschii Connectase) and known ligation enzymes (Sortase A, OaAEP1).
MmCET’s overall conformation: The apo form of MmCET exhibits a “jaw-like” conformation with a long groove rich in side chains; cysteine C65 and C193 form stable intermolecular disulfide bonds, locking the enzyme in a dimeric state, which restricts the crystal lattice (Figure S1D). By removing these cysteines through site-directed mutagenesis, the enzyme still exists in the same crystal form, and the crystallization process is unaffected. Structural analysis shows that the natural substrate recognition motif is not the “optimal” amino acid combination for MmCET, and the volume analysis of the substrate pocket of free MmCET suggests that better peptide substrates can be designed.
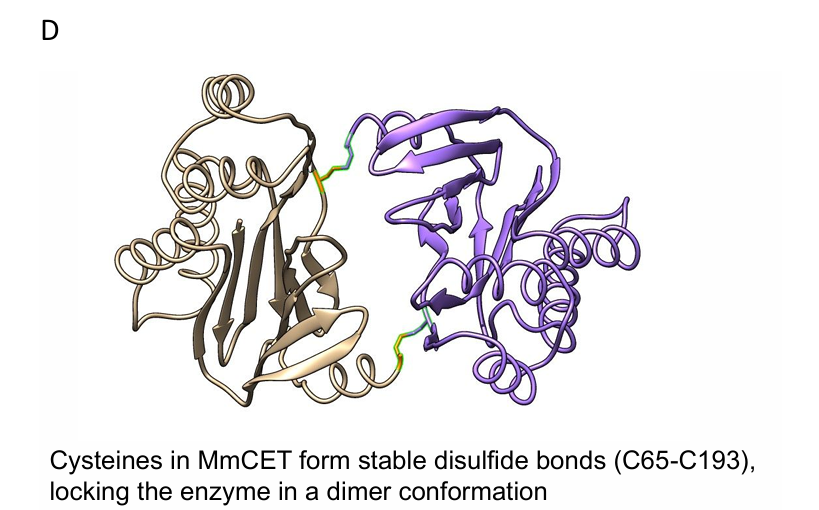 Figure S1D
Figure S1D
The crystal structure of the substrate-bound form: By synthesizing a series of peptide substrates and conducting peptide binding and ligation experiments (mass spectrometry monitoring of ligation efficiency), the study found that mutating F at the P4′ position of MmCET to V and D at the P7′ position to K (peptide sequence: RELASKD|PGAVDAKPLVVEI) significantly enhances ligation efficiency. Based on this, the structure of the binary complex formed by MmCET (T1A) with the substrate in the catalytic intermediate state was successfully resolved (PDB code: 8WKD), with a resolution of approximately 2.1 Å, revealing for the first time the fine interactions between MmCET and the preferred substrate.
Differences with MjCET and known ligation enzymes: Unlike the loose binding of substrates in MjCET, in the binary complex of MmCET, the entire peptide substrate is fully embedded in the substrate recognition groove: the P7-P1 positions (RELASKD) are accommodated in the C-terminal substrate pocket, the P1′-P3′ positions (PGA) are located in the catalytic pocket, and the P4′-P13′ positions (VDAKPLVVEI) perfectly fit the N-terminal substrate binding groove (Figure 1A), with the binding mode comparable to “a crocodile’s jaw biting a buffalo’s leg.” In contrast, Sortase A’s P2′-P5′ only loosely interacts with 2-3 surface residues (Figure 1D), recognizing sequences containing Gly-Gly (P1′-P2′) at the N-terminus, with no specificity for sequences beyond P3′, leading to easy re-recognition of ligation products and high reversibility; plant-derived AEP ligation enzymes like OaAEP1 lack a clear P1′-P5′ substrate recognition pocket, resulting in even less substrate recognition restriction (Figure 1D). In comparison, the extended N-terminal substrate recognition groove of MmCET provides great potential for specific engineering modifications.
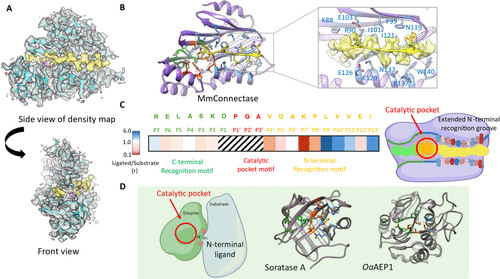
Figure 1
Furthermore, through a series of mutagenesis experiments, it was verified that the residues forming fine interactions with the substrate in the complex structure significantly affect the protein ligation activity of MmCET, confirming that this complex structure is indeed a “snapshot” of the catalytic intermediate.
2
Characteristics of the substrate binding groove of MmCET
The substrate binding groove of MmCET is divided into a C-terminal substrate pocket and an N-terminal substrate recognition groove, with significant differences in substrate recognition specificity, collectively determining the ligation precision of the enzyme.
C-terminal substrate pocket: Low specificity recognition: The interaction between the C-terminal substrate pocket of MmCET and the binding peptide is relatively weak. By performing continuous alanine scanning on the substrate P3-P6 positions (successively replacing each conserved recognition sequence amino acid with alanine) and measuring the catalytic efficiency using mass spectrometry to detect the product/substrate ratio, the results show that amino acid substitutions in this region have little effect on catalytic efficiency (Figure 1C), indicating that MmCET has a relatively relaxed sequence recognition for substrate P7-P2 positions.
N-terminal substrate recognition groove: High specificity recognition: The residues on both sides of the N-terminal substrate recognition groove form highly specific interactions with the substrate: residues 88-103, N119, I121, and H123 constitute the upper surface of the groove, while the side chains of α-helix 3 (residues 125-140) form the lower surface of the groove; the peptide substrate binding in this groove adopts a β-fold secondary structure, tightly binding to the residues on the upper and lower surfaces.
The residues facing upwards (P7′, P8′, P10′, P12′) interact with the residues on the upper surface of the groove: for example, F99 and I121 of MmCET bind to the P10′ position of the substrate, preferring small-sized hydrophobic residues; I101 and F123 bind to the P8′ position of the substrate, compatible with proline or medium-sized hydrophobic residues; E103 binds to the P7′ position of the substrate, preferring lysine (thus, when the P7′ position is replaced with alanine, the ligation efficiency significantly decreases).
The residues facing downwards interact with the residues on the lower surface of the groove: W140 and K137 of MmCET bind to the P11′ position of the substrate, repelling large-sized residues; N133 is close to the P9′ position of the substrate, preferring small-sized residues; the P4′-P5′ positions near the catalytic pocket bind to K88, compatible with medium-sized or negatively charged side chain amino acids.
The results of the alanine scanning experiments are entirely consistent with the above interaction characteristics, indicating that the “toothed” structure of the N-terminal groove only allows substrates with specific amino acid combinations to bind (Figures 1B, 1C), which is the core of MmCET’s high-precision recognition.
3
Relationship between the structure of MmCET’s catalytic pocket and ligation reversibility
By analyzing the MmCET-substrate complex structure, the study reveals the structural origins of substrate specificity and clarifies the influence of amino acid conformations in the catalytic pocket on the processivity of MmCET ligation.
Reaction mechanism of the catalytic pocket
In protease or ligation enzyme reactions, the C-terminal substrate must enter the catalytic pocket of MmCET. As shown in Figure 2A, the peptide bond after the aspartic acid (Asp) at the P1 position of the substrate can be attacked by Thr1 of MmCET (in the crystal structure as the T1A mutant, the enzyme is inactive), forming a catalytic intermediate— the C-terminal substrate is directly linked to Thr1 through the peptide bond, while the P1′ position proline (Pro) is cleaved. The energy advantage of this reaction arises from the instability of the peptide bond between Asp (P1) and Pro (P1′).
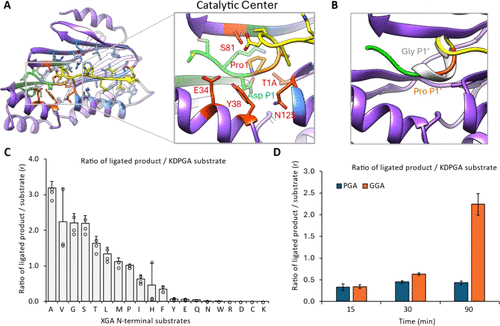
Figure 2
Structural observations reveal that the Pro-Gly-Ala sequence at the P1′-P3′ positions of the substrate is highly unstable due to bond angle restrictions (Figure 2B), which is a key reason for the ease of cleavage of this peptide bond.
Regulation of ligation processivity by P1′ position amino acids: To verify the hypothesis that “the conformational stability of the amino acid at the P1′ position affects ligation reversibility,” the study prepared 20 different peptide ligands with varying amino acids at the P1′ position, analyzing ligation efficiency through mass spectrometry to detect product/substrate ratios. The results show that when the P1′ position is occupied by small-sized amino acids such as glycine (Gly) or alanine (Ala), the peptide ligation efficiency is significantly enhanced (Figure 2C).
Using Coot software to dock the substrate model with Gly at the P1′ position reveals that this conformation forms a relatively stable secondary structure at the P1-P1′ ligation site, with minimal bond angle restrictions (Figure 2B). This indicates that if the P1′ position is occupied by small-sized amino acids, the peptide bond at the P1-P1′ ligation product is less likely to be cleaved again, avoiding reaction reversibility; whereas the natural Pro-Gly-Ala sequence is conformationally unstable, making the ligation product prone to re-cleavage and cycling as a substrate, leading to reaction reversibility.
4
Structure-based modification of the substrate recognition motif: Achieving irreversible protein ligation
To further verify the impact of the amino acid at the P1′ position on ligation processivity, the study designed time-dependent ligation experiments, using the product/substrate ratio as an indicator to compare the ligation efficiency of substrates with N-terminal Pro-Gly-Ala (PGA) and Gly-Gly-Ala (GGA) (Figure 2D).
GGA motif: Irreversible, high processivity ligation: The ligation processivity of Gly-Gly-Ala is significantly higher than that of Pro-Gly-Ala: when the N-terminal is GGA, the conformation of Gly-Gly-Ala in the catalytic pocket of the ligation product is stable, and the peptide bond between Asp (P1) and Gly (P1′) is stress-free, avoiding further cleavage; the reaction can continue, with substrates continuously consumed and products accumulating.
PGA motif: Reversible, low processivity ligation: When the N-terminal is PGA, the conformation of the residual Pro-Gly-Ala in the ligation product is unstable, making it easy for MmCET to re-cleave and cycle as a substrate; the reaction reaches equilibrium after 30 minutes, with no further changes in product and substrate concentrations.
This result indicates that by designing the N-terminal substrate recognition motif as GGA or other sequences compatible with the catalytic pocket conformation, the ligation reaction catalyzed by MmCET can be transformed into an irreversible reaction, significantly enhancing its processivity and overcoming the core limitation of natural MmCET, laying the foundation for its application as an efficient protein ligation enzyme.
5
Validation of MmCET in protein ligation experiments
Based on the conclusion that the GGA motif enhances peptide ligation processivity, the study further validated the practical application of MmCET using large, fully folded proteins as substrates and tested its compatibility in complex biological environments.
Efficient ligation of protein substrates
Using ubiquitin (Ub) as a model protein, we constructed Ub-KDPGA with a C-terminal containing KDPGA (P1′-P3′ as PGA) and GGA-Ub-mCherry with an N-terminal containing GGA; incubating 10 μM of both substrates with 2 μM of MmCET for 15 minutes resulted in a ligation efficiency exceeding 40% (estimated based on band intensity), which increased to over 60% after 60 minutes (Figure 3A), confirming that the combination of C-terminal KDPGA and N-terminal GGA can efficiently mediate the protein-protein ligation catalyzed by MmCET.
As a control, when the N-terminal substrate is PGA (i.e., the combination of Ub-KDPGA and PGA-Ub-mCherry), the ligation efficiency reaches a balance around 40%, with no further increase (Figure 3B), consistent with the previous results on the reversibility of peptide substrates.
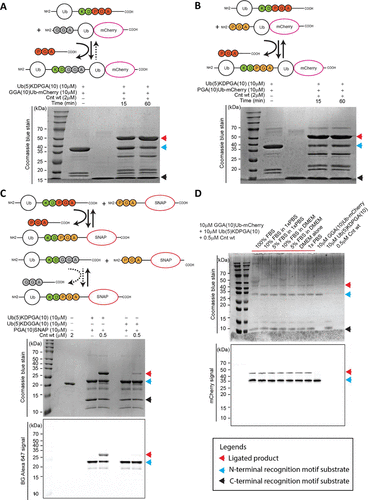
Verification of the cleavage resistance of the C-terminal GGA motif: To confirm that the irreversibility of the ligation product arises from the cleavage resistance of the C-terminal or internal GGA motif, the study constructed two ubiquitin carriers with C-terminal KDPGA and KDGGA, respectively, and conducted parallel ligation experiments with fluorescently labeled SNAP-N-terminal substrates (GGA-SNAP). The results showed that the product connected with the C-terminal KDGGA was almost not re-cleaved; whereas the product connected with the C-terminal KDPGA still exhibited some cleavage (Figure 3C), further confirming that the GGA motif can endow the ligation product with cleavage resistance, inhibiting reversible ligation.
Compatibility in complex biological environments: Unlike OaAEP1 (C247A), which requires a “clean” environment (serum GL residues or amino acids on cell surfaces may interfere with ligation), and Sortase A, which requires high concentrations (about 20 μM) to work, MmCET maintains high ligation activity even in complex environments containing fetal bovine serum (FBS): even under 100% FBS conditions, MmCET can still accurately recognize Ub-KDPGA and GGA-Ub-mCherry, achieving efficient ligation (Figure 3D). Additionally, in cell culture media containing FBS, using 0.05-2 μM MmCET and 0.5-20 μM GGA-SNAP, efficient and precise labeling of cell surfaces can be achieved, confirming the application potential of the structurally optimized MmCET in live cell environments.
 In AFM-SMFS studies, it is necessary to immobilize the target protein and construct protein polymers (polyprotein) to obtain characteristic “sawtooth” force-extension curves as reliable detection signals. The study combined MmCET with OaAEP1 (C247A) to achieve precise immobilization and stepwise polymerization of proteins, validating the application value of MmCET in single-molecule experiments.Precise immobilization of proteins: Using a “click chemistry + MmCET catalysis” strategy to immobilize ubiquitin (Ub): first, GGA (10)-N3 was modified onto DBCO-functionalized glass slides through click chemistry; then, under the catalysis of MmCET, GL-Ub-(5) KDPGA (10) was connected to the surface of the slides; to achieve AFM-SMFS detection, OaAEP1 catalyzed the connection of GL and NGL sequences, using Coh-NGL as a “pulling handle” connected to Ub; finally, using an AFM tip modified with GB1-XDoc, Ub was bound through the reversible interaction of Coh-XDoc. After stretching, the force-extension curve exhibited two characteristic unfolding peaks: corresponding to the contour length increment ΔLc≈24 nm for Ub, and ΔLc≈18 nm for GB1 (Figure 4B), confirming successful protein immobilization.Stepwise protein polymerization and single-molecule validation: Based on the above immobilization strategy, the study achieved stepwise tandem connection of proteins: after fixing Ub, without connecting Coh-NGL, GGA (10)-I27-NGL was directly connected to Ub through OaAEP1 catalysis, constructing (Ub)1-(I27)1; AFM detection showed a new characteristic unfolding peak for I27 (ΔLc≈28 nm).Repeating this stepwise enzymatic connection process, sequence-defined protein polymers were successfully constructed: (Ub)2-(I27)1, (Ub)2-(I27)2, (Ub)3-(I27)2; all polymers exhibited the expected unfolding peaks for Ub and I27 in AFM-SMFS detection (Figure 4C), confirming that MmCET can achieve orthogonal enzymatic connections with OaAEP1 (C247A), providing an efficient tool for the study of protein structure and function at the single-molecule level.
In AFM-SMFS studies, it is necessary to immobilize the target protein and construct protein polymers (polyprotein) to obtain characteristic “sawtooth” force-extension curves as reliable detection signals. The study combined MmCET with OaAEP1 (C247A) to achieve precise immobilization and stepwise polymerization of proteins, validating the application value of MmCET in single-molecule experiments.Precise immobilization of proteins: Using a “click chemistry + MmCET catalysis” strategy to immobilize ubiquitin (Ub): first, GGA (10)-N3 was modified onto DBCO-functionalized glass slides through click chemistry; then, under the catalysis of MmCET, GL-Ub-(5) KDPGA (10) was connected to the surface of the slides; to achieve AFM-SMFS detection, OaAEP1 catalyzed the connection of GL and NGL sequences, using Coh-NGL as a “pulling handle” connected to Ub; finally, using an AFM tip modified with GB1-XDoc, Ub was bound through the reversible interaction of Coh-XDoc. After stretching, the force-extension curve exhibited two characteristic unfolding peaks: corresponding to the contour length increment ΔLc≈24 nm for Ub, and ΔLc≈18 nm for GB1 (Figure 4B), confirming successful protein immobilization.Stepwise protein polymerization and single-molecule validation: Based on the above immobilization strategy, the study achieved stepwise tandem connection of proteins: after fixing Ub, without connecting Coh-NGL, GGA (10)-I27-NGL was directly connected to Ub through OaAEP1 catalysis, constructing (Ub)1-(I27)1; AFM detection showed a new characteristic unfolding peak for I27 (ΔLc≈28 nm).Repeating this stepwise enzymatic connection process, sequence-defined protein polymers were successfully constructed: (Ub)2-(I27)1, (Ub)2-(I27)2, (Ub)3-(I27)2; all polymers exhibited the expected unfolding peaks for Ub and I27 in AFM-SMFS detection (Figure 4C), confirming that MmCET can achieve orthogonal enzymatic connections with OaAEP1 (C247A), providing an efficient tool for the study of protein structure and function at the single-molecule level.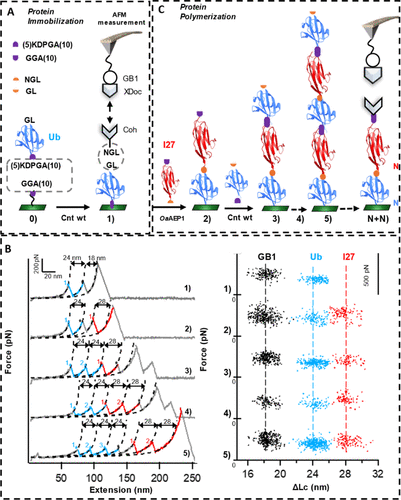 Figure 4
Figure 4
The discovery of ligation enzymes (Connectase) has added a class of enzymes with novel scaffolds to the protein ligation enzyme toolkit, opening new avenues for biochemical research and applications. Similar to plant asparagine endopeptidases (AEPs), this enzyme family was initially recognized for its protease activity. This study found that the key to transforming proteases into efficient ligation enzymes lies in the precise modification of key structural features. By elucidating the crystal structures of MmCET (the ligation enzyme from Methanococcus maripaludis) in both the substrate-bound state (in the catalytic transition state) and the unbound state (apo form), we revealed the core elements for achieving high-precision substrate recognition at the N-terminal motif.
Based on these newly discovered characteristics of MmCET, we successfully addressed a major limitation of this enzyme—enhancing its ligation processivity, and confirmed its excellent performance in tandem protein ligation through single-molecule experiments. This breakthrough not only highlights the application potential of ligation enzymes but also lays the foundation for applying similar “structure-based engineering modification strategies” to other protease scaffolds.
The authors designed “customized high-specificity protein ligation enzyme tools” to meet a wide range of biological application needs. MmCET possesses flexible structural features, especially its highly modifiable N-terminal recognition groove, providing unprecedented possibilities for specific engineering modifications and paving the way for achieving bioorthogonal ligation. A deeper understanding of the substrate recognition specificity of MmCET and related protein ligation enzymes will drive the development of the next generation of protein ligation enzymes, enabling them to achieve unparalleled precision and efficiency.