 Abstract:Lipid nanoparticles (LNPs) are lipid vesicles with a uniform lipid core that are widely used for the delivery of small molecule drugs and nucleic acids. They have recently gained significant attention due to their remarkable success as a delivery platform for COVID-19 mRNA vaccines. However, the application of mRNA-induced transient protein expression is not limited to the vaccine field for infectious diseases; it also provides new avenues for cancer vaccines, protein replacement therapies, and gene editing components for rare genetic diseases. However, naked mRNA itself is unstable and degrades rapidly due to nucleases and self-hydrolysis. mRNA encapsulated in LNPs can be protected from extracellular ribonucleases and aids in intracellular mRNA delivery.In this article, we will discuss the core features of LNPs used for RNA delivery. The focus will be on LNPs for mRNA delivery; however, examples of siRNA-LNP delivery will also be provided to highlight the commonalities and differences caused by nucleic acid structures.First, we will introduce the concept of LNPs, the advantages and disadvantages of using nucleic acids as therapeutic drugs, and the general rationale for the molecular composition of LNPs; we will also briefly introduce the latest successful experiences of LNP-based nucleic acid therapies in clinical settings. Second, we will elaborate on the theory and methods of LNP self-assembly. The common idea behind all preparation methods is to induce electrostatic interactions between nucleic acids and charged lipids, promoting nanoparticle growth through hydrophobic interactions. Third, we will decompose the components of LNPs based on their fundamental properties and uses, including recognized molecular design standards, commercial sources, the impact on intracellular transport, and contributions to LNP performance. One of the key components of LNPs is ionizable lipids that initiate electrostatic binding with endosomal membranes and facilitate cytoplasmic release; however, the role of other lipid components should not be overlooked as they are closely related to the stability, spacing, and distribution of LNPs. Fourth, we will review the structural properties of LNPs that significantly impact RNA delivery, including LNP size, charge, internal structure, lipid encapsulation, lipid membrane hydration, stability, and affinity for biomacromolecules; we will also discuss the detection techniques and regulatory methods for these properties.Finally, we will look ahead to the future of RNA therapies and address some issues that exist in the field of LNP formulation and optimization.
Abstract:Lipid nanoparticles (LNPs) are lipid vesicles with a uniform lipid core that are widely used for the delivery of small molecule drugs and nucleic acids. They have recently gained significant attention due to their remarkable success as a delivery platform for COVID-19 mRNA vaccines. However, the application of mRNA-induced transient protein expression is not limited to the vaccine field for infectious diseases; it also provides new avenues for cancer vaccines, protein replacement therapies, and gene editing components for rare genetic diseases. However, naked mRNA itself is unstable and degrades rapidly due to nucleases and self-hydrolysis. mRNA encapsulated in LNPs can be protected from extracellular ribonucleases and aids in intracellular mRNA delivery.In this article, we will discuss the core features of LNPs used for RNA delivery. The focus will be on LNPs for mRNA delivery; however, examples of siRNA-LNP delivery will also be provided to highlight the commonalities and differences caused by nucleic acid structures.First, we will introduce the concept of LNPs, the advantages and disadvantages of using nucleic acids as therapeutic drugs, and the general rationale for the molecular composition of LNPs; we will also briefly introduce the latest successful experiences of LNP-based nucleic acid therapies in clinical settings. Second, we will elaborate on the theory and methods of LNP self-assembly. The common idea behind all preparation methods is to induce electrostatic interactions between nucleic acids and charged lipids, promoting nanoparticle growth through hydrophobic interactions. Third, we will decompose the components of LNPs based on their fundamental properties and uses, including recognized molecular design standards, commercial sources, the impact on intracellular transport, and contributions to LNP performance. One of the key components of LNPs is ionizable lipids that initiate electrostatic binding with endosomal membranes and facilitate cytoplasmic release; however, the role of other lipid components should not be overlooked as they are closely related to the stability, spacing, and distribution of LNPs. Fourth, we will review the structural properties of LNPs that significantly impact RNA delivery, including LNP size, charge, internal structure, lipid encapsulation, lipid membrane hydration, stability, and affinity for biomacromolecules; we will also discuss the detection techniques and regulatory methods for these properties.Finally, we will look ahead to the future of RNA therapies and address some issues that exist in the field of LNP formulation and optimization.
1. Introduction
In recent years, lipid nanoparticles (LNPs) have demonstrated their practicality as delivery systems in RNA vaccines and therapies. Naked RNA is a negatively charged hydrophilic macromolecule that has difficulty entering cells due to electrostatic repulsion from the cell membrane and is rapidly degraded by ubiquitous ribonucleases (RNases). Therefore, it requires a protective layer to increase the chances of entering the interior of the cell. Since the cell membrane is primarily composed of lipids, using liposomes to encapsulate RNA can facilitate RNA’s passage through the cell membrane and its release into the cytoplasm. To achieve this, liposomes first require a positively charged lipid particle that can attach to the negatively charged RNA. However, liposomes composed of permanently cationic lipid particles can electrostatically disrupt the negatively charged cell membrane, leading to cytotoxicity. Therefore, the lipid structure is further improved to acquire positive charges through the acidic endosomal pathway; the LNP components also extend to structural lipids (which mimic cell membranes and shield positive charges) and poly(ethylene glycol)-anchored lipids (which prevent LNP aggregation and adverse reactions with the biological environment). The FDA’s approval of the first siRNA-LNP drug (Onpattro) and emergency use authorization (EUA) for mRNA-LNP COVID-19 vaccines (Comirnaty) and Moderna’s COVID-19 vaccine clearly indicates that LNP-based nucleic acid delivery methods are safe and suitable for various therapies. However, there is currently no universal solution suitable for all diseases, so the optimization of LNPs remains a significant challenge. This article will discuss the key scientific and manufacturing concepts required for selecting RNA-LNP delivery lipids.
2. Preparation
The preparation of LNPs relies on self-assembly, where lipid components spontaneously assemble into nanoparticles based on intermolecular interactions. The formation of LNPs begins with the electrostatic binding between negatively charged nucleic acids and positively charged lipids. Then, LNPs grow through hydrophobic and van der Waals interactions between lipid components. Due to the diversity of lipids, the uniqueness of nucleic acids, and the instantaneous nature of the mixing process, characterizing the early stages of self-assembly and their relevant effects on the final properties of LNPs remains highly challenging. Furthermore, LNP manufacturing protocols impact the self-assembly of the product in at least two aspects: the uniformity of LNPs and nucleic acid loading efficiency.
LNPs can be prepared using various methods, such as lipid film extrusion, lipid film rehydration, nanoprecipitation, and microfluidic mixing. However, the typical preparation method involves rapidly mixing water and lipid components. Microfluidic technology has become the preferred method for current preclinical research due to its good reproducibility. Advances in the manufacturing of novel microfluidic devices make this method easier to implement; high-throughput preparation of LNPs can also be achieved using other approaches, such as parallel microfluidic channels or improved traditional methods (e.g., pipette mixing and T-mixer). Generally, microfluidic mixing is beneficial for encapsulating hydrophilic components into the hydrophobic lipid core, but this process does not strictly require the participation of nucleic acids. Kulkarni et al. reported that as long as the lipid core is formed, LNPs can be assembled with siRNA in a T-mixer. Overall, the characteristics of the mixing method may affect the assembly efficiency and internal structure of LNPs, while the kinetic factors of the self-assembly process will determine the final nanostructure.
3. Formulation
3.1 Cationic and Ionizable Lipids
Cationic lipids (CLs) and ionizable lipids (ILs) initiate the first step of self-assembly through electrostatic interactions. Liposomes containing CLs are still widely used for nucleic acid delivery. However, due to toxicity issues and lack of in vivo efficacy, they have been replaced by pH-sensitive ILs. When formulated into LNPs, ILs are designed to exhibit electrical neutrality at physiological pH but become positively charged in acidic endosomal environments. This pH-responsive ionization enhances their efficacy while reducing toxicity, making them more suitable for nucleic acid delivery. These lipids typically comprise 30-50% of the total lipids in the formulation. Many studies are dedicated to fine-tuning the properties of ILs to further improve efficiency, especially in hard-to-reach tissues. The overall structure of CLs and ILs can be divided into three parts: (1) head group, (2) linker, and (3) tail (Figure 1).
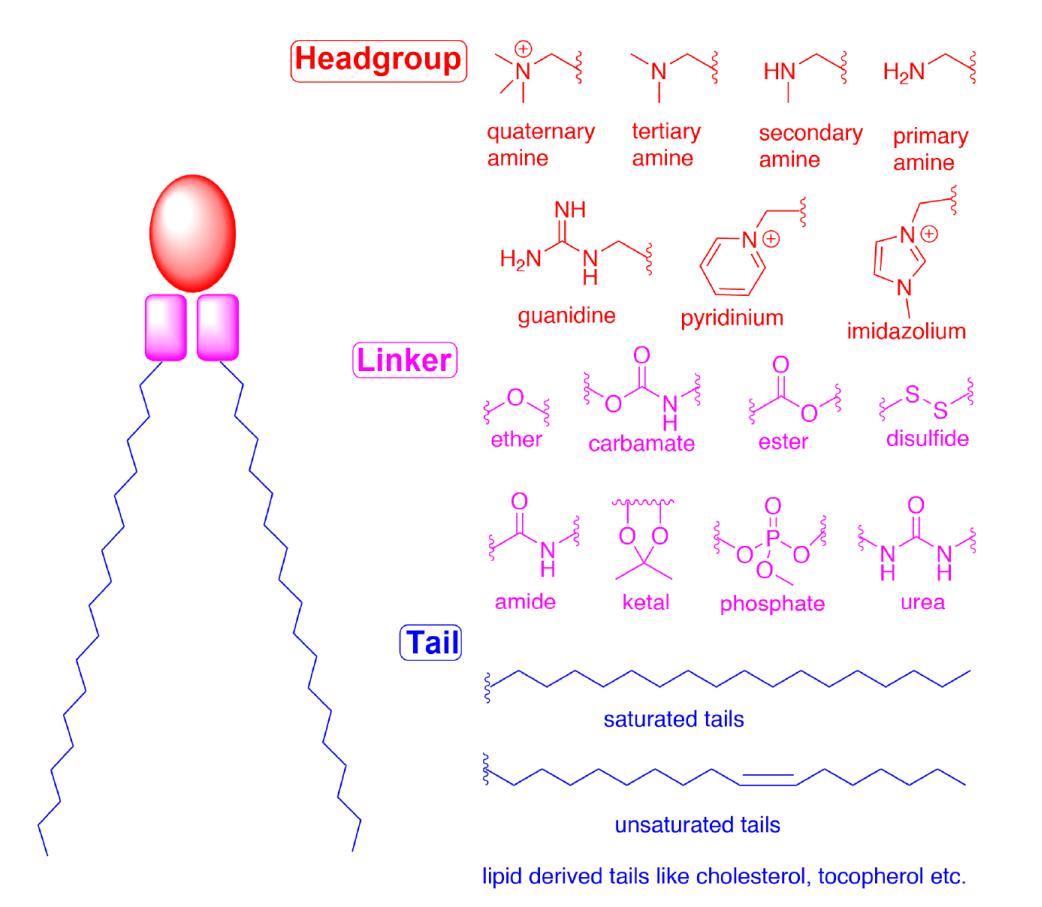
Figure 1 Schematic representation of the structures and components (head group, linker, and tail) of CLs and ILs
Head group The head group of ILs is usually positively charged. The size and charge density of the head group play an important role in nucleic acid encapsulation, LNP stability, interaction with cell membranes, and facilitating release in endosomes. ILs may also have multiple ionizable head groups, although typically only one is present. Typical groups include amines (from primary to quaternary), guanidines, and heterocyclic groups (see Figure 1). Clinical ILs (DLin-MC3-DMA, SM-102, and ALC-0315; see Figure 2) contain quaternary amine head groups with pH-responsive ionization. ALC-0315 and SM-102 head groups also contain terminal hydroxyl groups that can reduce the hydration of the head group and enhance its hydrogen bonding with nucleic acids, potentially improving transfection efficiency.
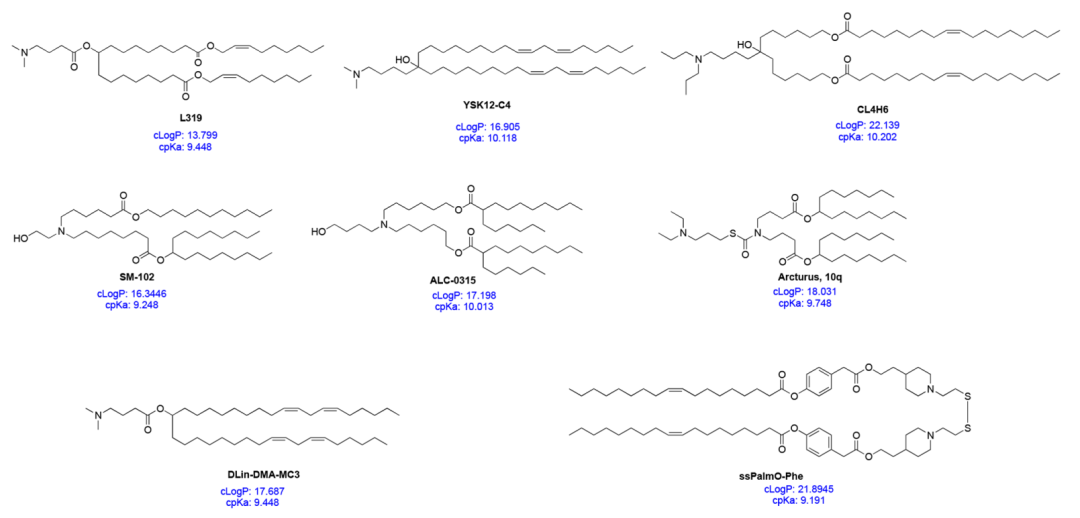
Figure 2 Specific IL structures and their cpKa and cLogP values
Linker The linker typically connects the head group to the tail and may also be included in the tail (SM-102 and ALC-0315; see Figure 2). Linkers affect the stability, biodegradability, cytotoxicity, and transfection efficiency of LNPs. Common linkers used in the design of CLs and ILs are shown in Figure 1. ILs may contain one or more linkers; however, for ease of synthesis, most ILs only contain one type of linker. Linkers can be classified as non-biodegradable (e.g., ethers and carbamates) and biodegradable (e.g., esters, amides, and thiols). Biodegradable linkers are preferred because they can be rapidly cleared in vivo, ensuring multiple administrations and reducing potential side effects. Notably, DLin-MC3-DMA, ALC-0315, and SM-102 all contain ester linkers. For SM-102, modifications around the ester group have been shown to affect the clearance, stability, and transfection efficiency of LNPs.
Tail The hydrophobic tail affects pKa, lipophilicity, fluidity, and fusogenicity, thereby influencing the formation and efficacy of nanoparticles. Typically, an IL contains 1 to 4 hydrophobic tails composed of 8 to 20 carbon atoms, either saturated or unsaturated. The degree of unsaturation has been shown to influence nucleic acid delivery by modulating membrane instability. DLin-MC3-DMA has two linoleic acid tails, while ALC-0315 and SM-102 contain two branched saturated tails presumed to be conical, aiding in destabilizing endosomal membranes and releasing nucleic acids into the cytosol.
ILs can be viewed as multicomponent molecules, where each part needs to be precisely designed to safely and efficiently encapsulate and deliver nucleic acids. Understanding the properties of ILs as a whole also aids in designing the next generation of ILs. One of these properties is the computed pKa (cpKa) of ILs, which can be easily determined in silico. The cpKa values of common ILs range from 9 to 10.5 (Figure 2). A recent study showed that cpKa is extremely close to the actual pKa of ILs. The cpKa of ILs seems to influence the overall pKa of the corresponding LNP formulation, such that when the cpKa of ILs is around 8.5−10.5, the pKa of LNPs is about 6−7. The difference between the cpKa of ILs and the pKa of LNPs appears to be fixed, approximately 2 – 4 units. Thus, cpKa can serve as a guiding tool for the design of new IL products. Two other less-discussed properties of ILs are cLogP and cLogD values, which represent the lipophilicity of the molecule in its non-ionized and ionized states, respectively. In a recent study, Rajappan et al. examined the effects of pKa, cpKa, and cLogD on the delivery of siRNA by LNPs and found that lipids with cLogD in the range of 10 – 14 were most effective. As some common ILs have cLogP values in the range of 15−20 (Figure 2), their lipophilicity should also be considered when designing the next generation of ILs. Since the degree of ionization (cpKa) and lipophilicity (cLogP) can affect the entire process from the initial formation of complexes with nucleic acids to the final nanoparticle formation and nucleic acid delivery, considering both parameters simultaneously before synthesizing ILs could greatly advance the discovery of efficient ILs. However, more research is needed to validate this perspective.
In addition to traditional CLs and ILs, there are examples of some zwitterionic lipids. In a recent study, Liu et al. synthesized a library of over 500 zwitterionic lipids named iPhos. These iPhos consist of an amine, a short hydrophobic tail, and a phosphate linker. The study suggests that the negatively charged phosphate group facilitates membrane fusion and leads to endosomal release. The optimal 9A1P9 formulations preferentially deliver the target nucleic acids to the liver and lungs.
In summary, the properties of each part of ILs influence the overall formulation and biological characteristics. Over the past 50 years, many systematic studies have been conducted to design ideal ILs. Some of these ILs have been approved by the FDA for the delivery of gene formulations. However, designing ILs that can efficiently and non-toxically deliver different types of gene formulations to non-hepatic targets still requires extensive research. Interested readers in IL synthesis can refer to an excellent related review.
3.2 Sterols
Cholesterol is a naturally abundant component of cell membranes and is one of the so-called structural lipids, frequently used in LNP formulations. Cholesterol typically constitutes 20-50% of the total lipids in LNPs. Cholesterol is abundant in animal cell membranes and can usually be extracted in large quantities from natural sources (e.g., wool). Despite its natural abundance, the role of cholesterol in cellular uptake has often been overlooked. Interestingly, substituting cholesterol with naturally occurring phytosterols, such as β-sitosterol (see Figure 3) and oxidized cholesterol derivatives, has been shown to significantly improve mRNA delivery, which may be due to specific lipid transfer mechanisms in endosomal pathways and enhanced endosomal release of LNPs. Although the reasons why LNPs containing β-sitosterol more easily evade the endosomal pathway are still unclear, their polymorphic shapes and multilayer organization may deform the endosomal membrane or prolong the duration of nucleic acid release. Cholesterol is primarily present in the shell of LNPs, which explains why modifications to sterol structures may induce changes in the surface organization of LNPs. Furthermore, a recent study indicated that when exposed to apolipoprotein E (ApoE), cholesterol migrates from the LNP core to the external lipid shell. These observations suggest that cholesterol and its derivatives may influence cellular recognition pathways, although cholesterol is relatively inert compared to ionizable lipids. However, there remain many unknowns and obstacles regarding potential modifications of cholesterol in LNP formulations. For example, phytosterols are not as abundant as cholesterol, and their scarce sources and high costs may hinder their application in LNP formulations. Similarly, the properties of raw materials and corresponding trace impurities may lead to potential batch differences in LNPs. The stereochemistry of sterols and lipids is an interesting topic worth exploring in LNP formulations. Does stereochemistry affect lipid recognition, transport, and circulation? Do enantiomers significantly influence lipid assembly on LNP surfaces? We hope to find answers to these questions soon.
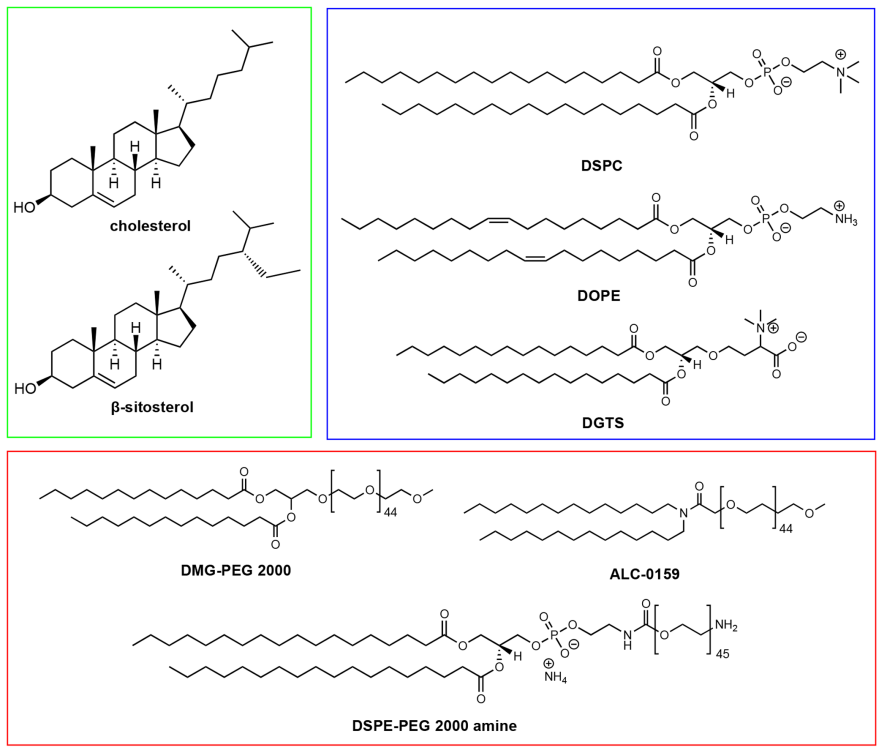
Figure 3 Examples of sterols (green), phospholipids (blue), and PEG lipids (red) used in LNP formulations
3.3 Phospholipids
Phospholipids assist in encapsulating nucleic acids and improving the stability of LNPs. Compared to other lipid components, their research is relatively less, typically constituting only 10 – 20% of the total lipids in the formulation. Phospholipids are used as structural lipids because they can spontaneously organize into lipid bilayers and have relatively high phase transition temperatures, ensuring the membrane stability of LNPs. Phospholipids are located on the periphery of LNPs, similar to cell membranes. These lipids are often semi-synthetic; for example, phosphatidylcholine typically comes from natural sources such as egg yolk and soybeans and can be chemically modified to include fatty acid tails. Distearoylphosphatidylcholine (DSPC) (see Figure 3) is a clinically approved structural lipid for LNPs, used in therapies such as siRNA (Onpattro) and mRNA vaccines against SARS-CoV-2. Structurally, DSPC consists of a phosphatidylcholine head group and two saturated 18-carbon tails, forming a tightly packed lipid bilayer. In LNPs, it is primarily located on the surface of nanoparticles, while a small amount is present in the nanoparticle core. Dioleoylphosphatidylethanolamine (DOPE) is another phospholipid frequently used in preclinical studies of LNPs. Due to its unsaturated tails, DOPE not only forms a more fluid lipid layer but also has an inherent ability to form hexagonal II (HII) phase structures. The HII phase structure is believed to facilitate membrane fusion between lipid membranes and endosomal membranes, thereby enabling the release of nucleic acid formulations into the cytosol. Multiple studies have shown that compared to DSPC, DOPE can enhance the transfection efficiency of RNA in phospholipid-based LNPs. Recently, Zhang et al. reported that DOPE can lead to the accumulation of C12-200 LNPs in the liver, while DSPC causes accumulation in the spleen upon intravenous administration, demonstrating the impact of structural lipids on the biodistribution of LNPs. We also found that replacing DSPC with natural glycosphingolipids in MC3-based LNPs affects mRNA transfection, with membrane lipids derived from plants—such as diacylglycerol trimethylhomoserine (DGTS) (see Figure 3)—exhibiting different transfection efficiencies depending on the route of administration. In summary, these studies highlight the importance of structural lipids in LNP-mediated RNA delivery.
3.4 PEG-Lipids
PEG-lipids (see Figure 3) are an important component for controlling the half-life of LNPs and cellular uptake. During the assembly of LNPs, the PEG chains are located in the nanoparticle shell due to their hydrophilicity and large size. Like other nanocarriers, PEG provides an external polymer layer to hinder the adsorption of serum proteins and mononuclear phagocyte systems, extending the circulation time in vivo. PEG also prevents the aggregation of nanoparticles during storage and in the bloodstream. Furthermore, the content of PEG lipids may determine the size of the particles. Another potential use of PEG lipids is to functionalize the surface of LNPs, making it possible for LNPs to bind with ligands or biomacromolecules. For example, Singh et al. targeted tumor therapy by conjugating hyaluronic acid to LNPs using DSPE-PEG-amine through NHS/EDC chemical conjugation, while Parhiz et al. conjugated antibodies to LNPs using DSPE-PEG-maleimide through SATA-maleimide chemical conjugation. Although PEG contributes to the stability and bioconjugation of LNPs, its desorption is also crucial for cellular transfection. The release of PEG from LNPs can trigger the conditioning effect of serum proteins (such as apolipoproteins and albumin), which is a key effector for receptor-mediated endocytosis of LNPs. Akinc et al. demonstrated that the binding of ApoE to LNPs leads to the internalization of low-density lipoprotein receptors (LDLRs) in hepatocytes. Since PEG lipids inhibit the binding of ApoE to LNPs, excessive PEG lipids can adversely affect cellular uptake and transfection of LNPs. LNPs with lower PEG lipid content exhibit higher acid delivery efficiency due to easier binding with ApoE. The length of the PEG lipid anchor is also an important factor determining the desorption rate. Mui et al. reported that the desorption of PEG from LNPs is inversely proportional to the length of the PEG-lipid anchor, as the hydrophobic interactions between PEG-lipid and LNP membranes increase with the length of the PEG-lipid anchor. Suzuki et al. proposed that the rate of PEG desorption may also influence the production of anti-PEG antibodies, and repeated administrations may lead to complications. The content of PEG lipids in intravenous LNP formulations rarely exceeds 2%; however, a dense polyethylene glycol layer may be beneficial for targeting extrahepatic sites. Lee et al. found that LNPs with 5% PEG lipid content accumulated more in tumors than those with 2.5% PEG lipid content, while Lokugamage et al. indicated that PEG lipids are essential for the delivery of nebulized LNPs. Therefore, the content and types of PEG lipids in LNPs may need to be carefully adjusted according to clinical needs.
4. Properties
The average size and size distribution of LNPs are important initial determinants of LNP quality and applicability in various applications, and these properties are typically studied using dynamic light scattering (DLS). Generally, the optimal size for LNPs is 20 – 200 nm, as this size allows them to be sufficiently stable in fluids (such as blood and lymph) while being able to pass through gaps. The size of LNPs is usually adjusted by changing the amount of PEG lipids or mixing parameters (such as flow rate and volume ratio). The size of LNPs may affect their endocytosis, biodistribution, degradation, and clearance, with different applications potentially requiring different particle sizes. For example, 45 nm siRNA LNPs are most effective for subcutaneous administration, while 80 nm siRNA LNPs are most effective for intravenous administration in mice. However, comparisons of various sizes of mRNA-LNPs in rodents and non-human primates indicate that non-human primates are less sensitive to the particle size of LNPs during intramuscular injection.
LNPs rely on surface charge to interact with cell membranes and the biological environment. Since cell membranes are negatively charged, negatively charged LNPs will be repelled by cell membranes and not taken up by cells. On the other hand, positively charged LNPs may directly disrupt cell membranes, causing cytotoxicity. This underscores the importance of ionizable lipids in LNP design: initially, LNPs containing ionizable lipids are neutral, avoiding any unnecessary electrostatic interactions, but become positively charged in acidic endosomal pH environments. The surface charge of LNPs is typically assessed using zeta potential measurements, a technique commonly used to evaluate colloidal aggregation. Although there is no strict classification, if the zeta potential falls between -20 and +20 mV, it is considered weakly charged. A common method to adjust the overall surface charge of LNPs is to modify the N/P ratio, or the ratio of ionizable lipids (N represents cationic amines) to nucleic acids (P represents anionic phosphates). Carrasco et al. suggested that increasing the N/P ratio of LNPs containing the ionizable lipid KC2 could increase surface charge and encapsulation efficiency. Interestingly, adding permanently charged lipids to LNPs may alter the preferential uptake of organs without increasing surface charge. Cheng et al. achieved selective organ targeting (SORT) in mice based on lipid charge: adding positively charged lipids to LNP formulations enabled preferential transfection of lung tissue, while negatively charged lipids directed transfection to the spleen.
Lipid encapsulation may affect many parameters, from membrane hydration and deformation ability to cellular uptake and nucleic acid release. A recent review summarized the basics of liposomes required for forming lipid vesicles. Simply put, each lipid can be described by an encapsulation parameter, which depends on the volume occupied by the lipid’s polar “head” and nonpolar “tail.” Structurally balanced lipids form cylindrical and lamellar phases, while “imbalanced” structures form hexagonal, cubic, and micellar phases. The inverted hexagonal phase (HII) exhibits the most significant membrane fusion-promoting effects. So far, the controlled preparation of non-lamellar phases in RNA-LNP delivery remains relatively rare, with nanocube liquid crystals being the most prominent example. However, LNPs may undergo structural changes when exposed to environmental stimuli. Heyes et al. used nuclear magnetic resonance spectroscopy (P NMR) to study the phase transition behavior of lipid particles containing a range of cationic lipids with different lipid tails, finding that lipids with lower temperatures for the lamellar to inverted hexagonal phase transition (TBH) exhibit better fusion-promoting capabilities, as demonstrated in gene silencing efficiency. Similarly, one of the membrane instability theories suggests that when ionizable lipids are exposed to the acidic pH of late endosomes, electrostatic interactions between ionizable lipids and phospholipids in endosomal membranes can lead to membrane rupture. Liu et al. recently reported evidence of inverted hexagonal phase formation when a novel ionizable lipid is exposed to endosomal mimetics, based on nuclear magnetic resonance spectroscopy. While pH-induced associations are the most common mechanism for lipid materials, other pathways leading to endosomal membrane instability have been discussed in our recent review.
Due to differences in lipid phases and overall polarity, lipid membranes may capture water, thereby altering membrane fluidity or deformability, which may affect lipid membrane fusion. Membrane hydration may also influence the potential response to pH, which is often an important environmental trigger for nucleic acid release. The pH fluctuations involved in the lifecycle of LNPs are illustrated in Figure 4. When LNPs enter intracellular spaces, they are trapped in endosomes, which gradually acidify as they mature into lysosomes. Therefore, higher hydration levels within the lipid membrane may affect the acidification kinetics and accelerate membrane destabilization. Koitabashi et al. studied the impact of pH on membrane stability in siRNA-LNPs through Laurdan experiments, finding that membrane hydration correlates positively with gene silencing efficiency; however, this study did not focus on acidification kinetics. An interesting phenomenon regarding membrane hydration is that nuclear magnetic resonance results indicated that siRNA-LNPs have less water content than mRNA-LNPs of the same formulation, which may be due to the longer length of the hydrophilic RNA chain. Carrasco et al. further demonstrated these points, finding that LNPs with lower N/P ratios contained more mRNA and lipids in individual nanoparticles and had higher dielectric constants, suggesting that LNPs with low N/P ratios are more easily hydrated than those with high N/P ratios. A higher RNA loading also improves transfection. Therefore, mRNA-LNPs may be more sensitive to environmental changes, although changes in pH sensitivity may not correlate with the timescale of biological processes. Further exposure to the biological environment complicates the issue of LNP shell hydration.
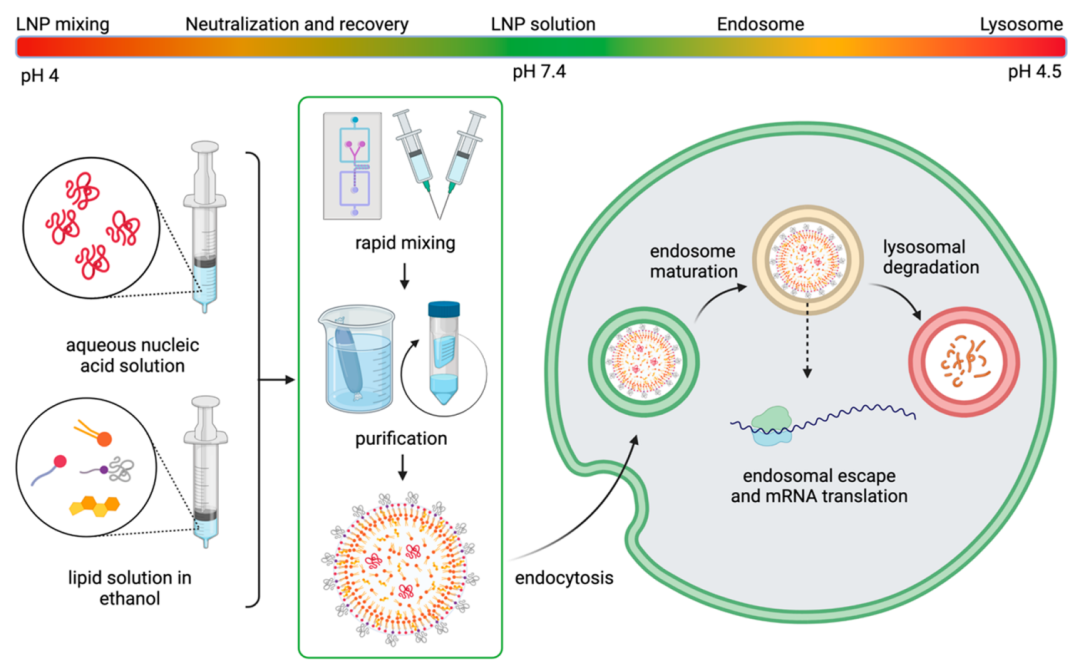
Figure 4 pH changes involved in the lifecycle of LNPs
The inherent aqueous environment of LNPs also poses a threat to their long-term stability. Pure nucleic acids can rapidly deteriorate under environmental conditions due to exogenous RNase degradation or self-hydrolysis. While LNPs can protect nucleic acids from enzymatic degradation, they are prone to aggregation due to thermodynamic factors (e.g., minimizing phase separation), which may lead to the loss of nucleic acids during nanoparticle fusion, ultimately affecting transfection efficiency. Low-temperature storage and lyophilization can protect RNA; however, ice crystallization can damage LNPs, although the addition of cryoprotectants such as sucrose seems to alleviate this issue. Interestingly, LNPs may alter the preferential uptake of organs based on storage conditions, which may be a result of their reorganization. From a practical perspective, the recently developed mRNA-LNP vaccines for COVID-19 have created a logistical infrastructure for RNA therapies, which may alleviate many concerns regarding LNP stability, storage, and transportation. These vaccines exhibit long-term stability at frozen temperatures (-20°C) for up to 6 months and at room temperature for up to 30 days, greatly facilitating the accessibility of these breakthrough therapies. Notably, the Pfizer/BioNTech and Moderna vaccines have different storage requirements, indicating that variations in LNP formulations may significantly alter their nucleic acid affinity and LNP stability. As there is currently no established method for accelerated stability testing of these amorphous materials, it means that LNP stability can only be empirically assessed at discrete time points, and methods like differential scanning calorimetry (DSC) may provide valuable insights into LNP degradation. Finally, predicting the downstream properties of LNP-based RNA therapies requires a deeper understanding of the self-assembly process of LNPs.
The internal organization of lipid nanoparticles remains a universally unresolved issue. Some studies suggest that siRNA-LNPs have a “sandwich” structure, with siRNA linked to the lipid layer on the outside of LNPs. In contrast, mRNA may extend along the periphery of LNPs, occupying the water-lipid interface within “bubble-like” LNPs, or occupying interconnecting water channels that mRNA may be displaced from due to its hydrophilicity. mRNA may also dissociate from the lipid core into the aqueous cavity. Our recent research utilizing cryo-transmission electron microscopy indicates that any deviation from uniform spherical LNPs (e.g., polymorphic shapes, multilayer structures, or aqueous cavities) may enhance mRNA transfection efficiency; however, it remains unclear whether the molecular recognition of lipid components will influence the enhancement of transfection. Changes in lipid structures may also be affected by solubility. While lipids are typically dissolved by heating or sonication before mixing, Yanez Arteta et al. detected 2D crystals of cholesterol within LNP lipid membranes using small-angle X-ray scattering. Lipid organization may also change when exposed to biological environments, as detected by small-angle neutron scattering experiments, revealing that LNPs undergo rearrangements on the surface and core after exposure to ApoE (a protein abundant in serum). The uncertainties regarding lipid structures related to RNA delivery require further research.
Nanoparticles are prone to nonspecific adsorption of proteins, forming a biomolecular corona at the interface. The formation of biomolecular coronas on nanoparticles alters their surface properties and physicochemical characteristics, significantly impacting their biodistribution and endocytosis. ApoE, as a component of the biomolecular corona, is recognized to influence LNP-mediated nucleic acid delivery. ApoE is widely present in the liver, primarily synthesized by hepatocytes, which is partly why intravenously injected LNPs often enter the liver. ApoE is inherently involved in cholesterol metabolism, facilitating receptor-mediated endocytosis by forming lipid complexes and transporting them to expressed low-density lipoprotein receptors (LDLRs). This concept also applies to LNPs. Akinc et al. found that in the absence of ApoE or LDLR, the cellular uptake of LNPs containing the ionizable lipid KC2 was significantly reduced compared to normal conditions. Dong et al. further supported this notion, reporting that LNPs containing the ionizable lipid cKK-E12 were more effectively internalized in the presence of ApoE. Other lipids in LNP formulations may also associate with ApoE bound to LNPs. Zhang et al. reported that when other lipid components in LNPs are the same, DOPE interacts more strongly with ApoE than DSPC, indicating that changes in phospholipid structures alter the affinity of ApoE for LNPs. Sebastiani et al. reported that the adsorption of ApoE on mRNA-LNPs leads to the rearrangement of lipid components within the nanoparticles, with some cholesterol migrating from the core of the nanoparticles to the shell after exposure to ApoE. Kim et al. reported that changes in PEG content and the binding of PEG lipids to mannosides can control the cell-specific delivery of mRNA in the liver. Interestingly, astrocytes in the brain and central nervous system can also produce substantial amounts of ApoE. Tanaka et al. studied the delivery of mRNA-LNPs via injection into the cerebral ventricles of mice, finding that astrocytes successfully absorbed ssPalm LNPs. However, ApoE is not the only known protein associated with differences in cellular uptake. Recently, Miao et al. confirmed that serum albumin allows LNPs to enter cells through non-ApoE pathways, highlighting the importance of other serum proteins. Despite these intriguing perspectives, it remains unclear how to fully harness the potential of biomolecular coronas. The biomolecular corona and LNPs are related to immune system recognition, blood circulation time, and biodistribution, and finding ways to manipulate biomolecular coronas could enable specific uptake by cells, tissues, or organs.
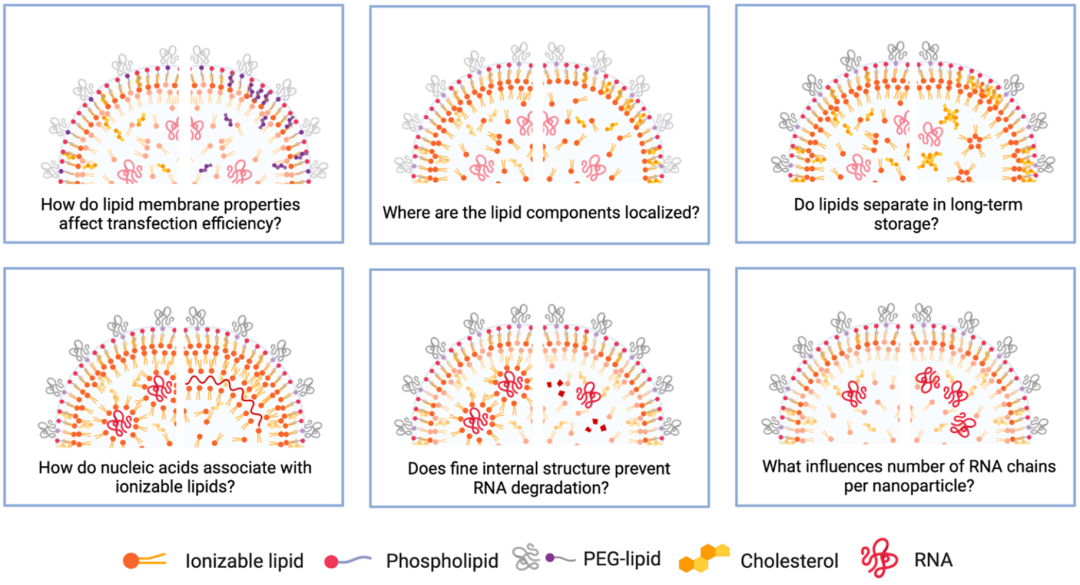
Figure 5 Unresolved issues regarding LNP structures
5. Conclusion and Outlook
LNPs are highly customizable nucleic acid carriers that show great potential in mRNA vaccines. We should not overlook their potential value in treating rare diseases and cancer. mRNA therapies can help produce therapeutic proteins to restore the function of damaged tissues or organs. A wealth of scientific research is underway globally to design and refine individual components of LNPs for the efficient and safe delivery of nucleic acids of interest. However, the science of LNPs is still in its infancy, with many unresolved questions, some of which are listed in Figure 5. Undoubtedly, the ongoing public interest in mRNA vaccines will inspire research efforts in this field, and we cautiously optimistic that we are witnessing a new era of nanomedicine.
Original source: Chemistry of Lipid Nanoparticles for RNA Delivery. Acc Chem Res. 2022 Jan 4;55(1):2-12.
