
Soybeans are an important source of plant protein and oil crops. According to their sedimentation coefficients at an ionic strength of 0.5 mol/L, they can be divided into four components: 2S, 7S, 11S, and 15S globulins. Among them, 7S and 11S globulins are the main storage proteins, accounting for about 80% of the total protein. In addition, recent years have seen the discovery of a membrane protein—lipoprotein (LP)—wrapped around the oil bodies in soybean. This component is a complex formed by the binding of soybean globulin, β-conglycinin, and oil body proteins with phospholipids. The structure and functionality of soybean protein components significantly affect the sensory quality of soy products such as tofu, soy milk, and plant-based meat.
Professor Zhu Xiuqing and others from the School of Food Engineering at Harbin University of Commerce review the current research status of soybean protein component structures, focusing on the preparation processes of soybean protein components and summarizing the functionalities of different soybean protein components, aiming to provide references for the quality control of soybean or soybean protein products.

Structure of Soy Protein Components
11S globulin is the most abundant globulin in soybeans, mainly consisting of soybean globulin rich in sulfur-containing amino acids, with a molecular weight ranging from 320 to 375 kDa. Its structure under neutral conditions is a hollow hexamer composed of five types of subunits, with each subunit formed by an acidic polypeptide and a basic polypeptide linked by disulfide bonds. The isoelectric point is approximately pH 5.8. The 7S globulin mainly consists of β-conglycinin, γ-conglycinin, and basic 7S globulin. Among them, β-conglycinin is the main component of 7S globulin, with a molecular weight of about 180-210 kDa, existing in a trimeric form under neutral conditions, formed by the aggregation of three subunits: α, α′, and β. The isoelectric points of α and α′ subunits are 5.2 and 5.3, respectively, while the β subunit consists of four components (β1-β4) with an isoelectric point ranging from 5.8 to 6.2. The structure of β-conglycinin is particularly special, as the α and α′ subunits have extended regions in addition to the core region, and all three subunits are N-glycosylated, meaning that the N-terminal of all three subunits is linked to high-mannose polysaccharides, which makes them distinctly different in solubility from soybean globulin. The denaturation temperatures of 11S and 7S globulins also vary in different experiments, generally with β-conglycinin denaturing at 68-72 °C and soybean globulin at 86-90 °C. The content ratio of 11S to 7S globulin in soybean grains usually ranges from 0.5 to 1.3, which can be attributed to differences in growth environment and genotype.
Additionally, the 15S globulin content in soybean protein is the least, approximately 5%, but gel filtration chromatography analysis indicates that 15S globulin can be viewed as a dimer composed of 11S globulin subunits. The 2S globulin mainly consists of protease inhibitors and other anti-nutritional factors, and although its content is only 8%, it can trigger respiratory allergic reactions in humans, hence it must be removed during processing. LP contains about 70% protein and 10% lipids, mainly consisting of lipoxygenase and membrane proteins. The proteins can be divided based on molecular weight into oil body proteins (24, 18 kDa), proteins derived from vacuoles (34 kDa), as well as subunits and peptides from 7S and 11S globulins, with the subunits of 7S and 11S globulins accounting for about 60% of the total protein content in LP. The proteins in LP show low sensitivity to Coomassie Brilliant Blue staining, making it difficult to roughly quantify their protein composition using sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE).
Preparation of Soy Protein Components
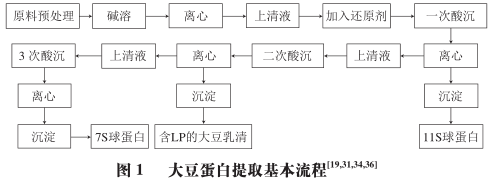
Currently, common soybean protein products include defatted soybean flour, soybean concentrate, and SPI. Among them, SPI has the highest protein content, reaching over 90%; soybean concentrate follows with about 70%; while defatted soybean flour only removes most of the lipids from the raw materials, retaining about 50% of polysaccharides and other non-protein components, but it is the raw material for SPI. They all have complete production process lines. However, the extraction methods for 7S, 11S globulins, and LP have not yet reached production line standards.
Nagano et al. proposed a purification scheme for 7S soybean globulin for the first time to determine the dynamic rheological parameters of 7S soybean globulin gel. The optimal conditions for separating 7S globulin are considered to be 0.25 mol/L NaCl, pH 5.0, and 4 °C, while sodium bisulfite (SBS) acts as a reducing agent, thereby increasing protein extraction rates. This method is the basic separation scheme for the “three-step acid precipitation” of soybean protein.
Since 1992, numerous studies have focused on optimizing the extraction efficiency of soybean protein, mainly based on the effects of process parameters such as temperature, pH, and ionic strength on protein conformation during extraction.
In fact, SPI not only contains 7S and 11S globulins, but also a large number of membrane proteins surrounding the protein bodies and oil bodies in the grains. Studies have shown that soybean protein extracted using traditional alkaline solubilization and acid precipitation methods has poor taste and still retains some proteins bound with lipids, namely LP. The discovery of LP indirectly explains the impurity and low yield of 11S and 7S globulins extracted by previous methods, and it has also prompted some researchers to begin exploring its functionality.
The extraction processes and parameters of soybean protein components in different studies are shown in Figure 1 and Table 1.
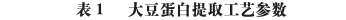

Functionality of Soy Protein Components
Processing conditions can lead to the transformation of proteins from their native state to an intermediate state and finally to complete denaturation. Thus, the natural state of proteins does not entirely determine their functionality; it is also necessary to understand the structural transition behaviors of proteins during folding and unfolding in environments such as solutions, interfaces, and gels.
The solubility of proteins is usually the primary characteristic to be examined for the production of gels, emulsions, beverages, and other products, which can be expressed as the proportion of nitrogen content in a specific solution to the total nitrogen content of the sample. Solubility is not only related to the amino acid composition, sequence, molecular weight, and protein conformation but also to environmental factors such as ionic strength, pH, and temperature. Based on the protein structure, it can be inferred that the polysaccharides linked to the N-terminus of β-conglycinin increase its solubility in water under neutral conditions. Research has shown that the solubility of 7S globulin is maximized at pH 7.0 (90%), followed by SPI (80%) and 11S globulin (60%). The solubility curves of the three proteins change in a U-shape with pH, with both 7S globulin and SPI having the lowest solubility around pH 4.5, while 11S globulin reaches its lowest solubility at pH 5.0, indicating that the isoelectric points of 11S and 7S globulins are approximately 4.5 and 5.0, respectively. Additionally, as the salt concentration in the dispersion increases, the sensitivity of the solubility of the three proteins to pH decreases.
The solubility of soybean protein is also affected by thermal aggregation behavior. Some researchers have proposed the Lumry-Eyring nucleation aggregation model, which suggests that protein aggregation consists of multiple stages, including conformational changes, pre-nucleation, irreversible aggregate nucleation, increased aggregation, and self-association of aggregates. Studies have shown that after preheating at 80 °C, SPI only exhibits an endothermic peak corresponding to 11S globulin at 97.6 °C, which may be due to the complete denaturation and unfolding of 7S globulin at 80 °C, forming more stable aggregates with 11S globulin. The aggregation behavior of β-conglycinin and soybean globulin during heating is different. The solubility of β-conglycinin remains relatively unchanged during heating from 50 °C to 100 °C, while the solubility of soybean globulin decreases as the temperature increases. When β-conglycinin and soybean globulin are mixed in ratios of 4:1, 2:1, and 1:1, the addition of β-conglycinin leads to smaller aggregate sizes and increased solubility. Hydrophobic interactions may be the primary driving force for protein aggregation. The thermal aggregation mechanisms of the two proteins are shown in Figure 2. When aggregation occurs, for β-conglycinin, once the hydrophobic residues are covered and aggregates are formed, the polysaccharides and hydrophilic groups on the surface provide repulsive forces to inhibit other protein monomers from approaching; however, soybean globulin’s basic polypeptides contain more hydrophobic amino acids, so upon unfolding, more active sites are exposed. Although some active sites are covered during aggregation, hydrophobic residues still exist on the surface of the aggregates, leading to continued aggregation of active sites; when the two proteins coexist, the surface of the globulin aggregates is no longer covered by hydrophobic residues, but rather by the hydrophilic groups of β-conglycinin, thus terminating the aggregation behavior. Furthermore, this thermal aggregation behavior between proteins is also concentration-dependent. Increasing protein concentration decreases the distance between proteins, effectively promoting aggregation.
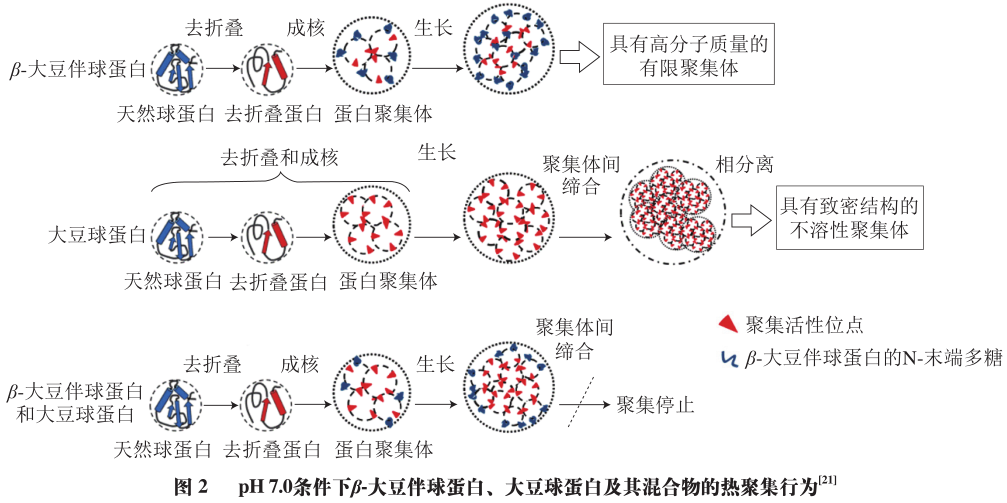
Improvements in breeding and protein extraction processes have provided better methods for obtaining high-purity soybean proteins. Using low-heat-treated soybean meal as raw material, high-purity β-conglycinin subunits were successfully obtained through DEAE-Sepharose fast flow chromatography combined with immobilized metal ion affinity chromatography, with β subunit content at 91.4%, α subunit content at 95.0%, and α′ subunit content at 92.1%. Based on this, it was found that the thermal aggregation of 7S globulin is mainly dominated by the β subunit, while the thermal aggregation of 11S globulin is dominated by the basic polypeptides. During heating from 50 to 90 °C, the molecular weight of the component corresponding to the chromatographic peak significantly increases for the β subunit (greater than 669 kDa), while the molecular weight distribution of the α and α′ subunits remains stable at around 29 nm under different temperatures. The Z-average diameter of the β subunit reaches 70 nm during heating at 50 °C and increases to 158 nm at 90 °C, indicating that the β subunit in 7S globulin is more prone to aggregation. Comparing the surface hydrophobicity of different protein subunits and polypeptides shows that the surface hydrophobicity of 11S globulin is approximately 5,000 at 60 °C, sharply increasing with temperature, peaking at 12,000 at 80 °C; the hydrophobicity of acidic polypeptides changes little during heating, also peaking at 4,200 at 80 °C. The peak values of surface hydrophobicity for 7S globulin, β subunit, and α and α′ subunits fluctuate around 8,000, 7,800, and 6,000, respectively. Additionally, the surface hydrophobicity of proteins, subunits, and polypeptides increases rapidly within the first 10 minutes of heating. This indicates that globulin molecules first undergo unfolding, and after being heated for a certain time at the denaturation temperature, the globular structure unfolds, exposing more hydrophobic residues and increasing surface hydrophobicity. The differences in thermal aggregation behavior between these subunits and polypeptides may be related to the amino acid composition of the primary structure of the proteins, for instance, basic polypeptides containing a high proportion of hydrophobic amino acids such as Val, Leu, and Ala can form insoluble aggregates at room temperature, and the β subunit also contains many hydrophobic amino acids, with only one N-linked high-mannose polysaccharide in its structure, while the α and α′ subunits each contain two.
Gelation is a special state of matter that lies between solid and liquid, and most foods can be consumed in gel form. When the intermolecular interactions of protein sols increase and cross-linking increases to a certain extent, they transform into gels. The basic formation process of thermally induced soybean protein gels is as follows: soybean protein disperses in water, initially forming a sol in a tightly coiled form; as the temperature rises, the proteins gradually denature and unfold, neighboring molecules first form aggregates, while high temperatures intensify molecular motion, leading to more frequent hydrophobic interactions between proteins. Once equilibrium is achieved, a gel with a certain network structure is formed, with some free water trapped within the gel network. In this formation process, covalent interactions via disulfide bonds play a dominant role, alongside non-covalent interactions such as hydrogen bonds and electrostatic repulsion. The critical protein concentrations for gel formation of 11S and 7S globulins in 35 mmol/L phosphate buffer (pH 7.6) at 100 °C are 2.5% and 7.5%, respectively. The thermal gel of 11S globulin primarily forms through disulfide bonds and electrostatic interactions, while the thermal gel of 7S globulin forms through hydrogen bonds. Therefore, pH and temperature can also influence the structure and properties of gels by affecting the aggregation of proteins.
Based on the higher aggregation degree of soybean globulin after heating, it can be inferred that its storage modulus (G′) for thermally induced gels should also be relatively large. The temperature scanning results of thermally induced gels made from the same soybean, with 95% purity for soybean globulin and 60% purity for β-conglycinin, indicate that at pH 3.8, the G′ of soybean globulin and β-conglycinin gels at the end of cooling are 4,000 Pa and 2,500 Pa, respectively, while at pH 7.6, the G′ of the former is 7,400 Pa, and that of the latter is 5,400 Pa. The breaking stress of soybean globulin gels at pH 3.8 and pH 7.6 is also significantly greater than that of β-conglycinin gels, at 46.2 kPa and 18.1 kPa for the former, and 2.1 kPa and 2.2 kPa for the latter, respectively.
The differences in G′ between these two protein gels may also be due to differences in sulfur-containing amino acid content. The methionine and cysteine content per unit of protein in 11S globulin is 3-4 times that of 7S globulin, thus 11S globulin has a stronger cross-linking network during gel formation. Studies have shown that the G′ of thermally induced gels is not directly related to the ratio of 11S to 7S globulins (r < 0.50) but depends on the content of the A3 acidic subunit, which has a relatively low cysteine residue content. Dynamic rheological analysis results of SPI thermally induced gels made from 11 different soybean globulin and β-conglycinin subunit-deficient soybeans show that when the A3 subunit accounts for less than 2% of the total protein content, there is a correlation (r > 0.967) between the content of 11S globulin subunits and the G′ when the gel is fully formed; when the A3 subunit accounts for more than 2% of the total protein content, there is no correlation between the G′ when the gel is fully formed and the composition of soybean proteins, and there is no significant difference in the G′ of gels from varieties with high or low 11S globulin content, indicating that 7S globulin is the main structural monomer of thermally induced gels, which may be due to the insufficient unfolding of 11S globulin at 90 °C. Furthermore, since protein unfolding is a prerequisite for gel formation, it can be predicted that the gel formation temperature of β-conglycinin, which has a lower thermal transition temperature, is lower than that of soybean globulin. The gel formation temperature for varieties with higher 7S globulin content is 74.2-82.2 °C, while the gel formation temperature for varieties with higher 11S globulin content ranges from 86.2 to 90.2 °C.
3.3 Rheological Properties
Rheological testing can reflect the phase transition of materials during processing to some extent by applying large shear forces or shear frequencies to disrupt the material structure, specifically characterized by parameters such as viscosity, G′, G′′, and torque. For high-moisture extruded soybean protein products, the velocity gradient caused by different viscosities is key to forming their fibrous structure. Steady-state scanning results of four SPI gels with similar sources and protein content (over 90% of dry matter) indicate that under the same conditions, the complex viscosity of SPI gels after 60s of treatment is 44 kPa·s (SPI 1), 43 kPa·s (SPI 2), 34 kPa·s (SPI 3), and 14 kPa·s (SPI 4), with this difference attributed to varying degrees of denaturation caused by different extraction processes. The protein content of soybean concentrate is about 67% of the total dry matter, but under the same conditions, its viscosity (101 kPa·s) is more than twice that of SPI viscosity. This indicates that a higher proportion of polysaccharides and other components in the raw materials also affects viscosity.
Most studies discuss the impact of the physicochemical properties of protein components on gel structure formation, but this is insufficient to explain the changing behavior during gel processing. The curvature and connection modes of chains formed by unfolded proteins determine how the gel will deform at the microscopic level, for example, gels with curved chains mainly deform by bending, while linear and interconnected chains deform by stretching. Based on this, studies have shown that the more curved the protein chain, the greater the strain at which the gel breaks, as the curved chain must first straighten before breaking. Through large amplitude oscillatory shear experiments, it was found that SPI, high 11S protein (11S globulin content 72.1%, 7S globulin content 3.3%), and high 7S protein (7S globulin content 30.4%, 11S globulin content 4.2%) have different inherent viscosities and intermolecular interaction strengths when forming thermally induced gels. Under low protein content (6%) conditions, the G′ of the three is ranked as follows: high 11S protein (600 Pa) > SPI (300 Pa) > high 7S protein (60 Pa). The G′ at low protein content reflects the inherent viscosity of the dispersed phase when molecular collisions, aggregation, and other interactions do not occur or occur less frequently, thus high 11S protein has higher inherent viscosity, followed by SPI. However, high 7S protein exhibits higher intermolecular interactions, which can be reflected in the G′-protein concentration curves of the three proteins, where the curve for high 7S protein has the steepest slope (281.25), followed by SPI (266.67), and high 11S protein (150), indicating that with each unit increase in protein concentration, the molecular collisions and aggregation of high 7S protein are greater, and G′ increases faster. This may be due to the insufficient unfolding of soybean globulin, where the rigid structure of globulin particles inhibits intermolecular cross-linking. For high 7S protein gels, the strong flexibility of β-conglycinin leads to more branched particles, with tighter interconnections, such that as strain increases, the initially dissociated particles intertwine to form new clusters. Observations using confocal laser scanning microscopy and scanning electron microscopy reveal that SPI and high 11S gels have relatively dense networks composed of larger aggregates, while high 7S protein gels are rougher and more uneven, with smaller and more branched particles.
Although soybeans have many applications in food production in China, due to the diversity of their components and extraction processes, companies still rely heavily on experience to control product quality. The correlation between raw material composition, structure, functionality, and sensory quality remains relatively vague. This article introduces recent knowledge related to the composition of soybean protein and industrial production, focusing mainly on protein extraction processes and functionality. There are still gaps in the separation identification of various soybean components and the characterization methods for protein functionality. Furthermore, to achieve green production and reduce carbon emissions, the utilization of by-products such as okara and soybean meal should be encouraged, for example, using extrusion cooking to produce plant-based meat products, developing soybean bioactive peptides, and producing biobased plastic products.
This article, “Research Progress on Soy Protein Component Preparation and Functionality,” is sourced from “Food Science” Volume 44, Issue 13, pages 360-367, 2023, by authors Luan Binyu, Zhang Bo, Meng Ang, et al. DOI:10.7506/spkx1002-6630-20220623-256. Click below to read the original text for more information.

Intern Editor: Mei Juan; Editor: Zhang Ruimei. Click below to read the original text to view the full text. Images are sourced from the original article and Shutterstock.
Recent Research Highlights
“Food Science”: Professor Gao Ruichang et al. from Jiangsu University: Monitoring Freshness of Fish Meat Using High-Sensitivity Indicator Films and Mechanism Analysis
“Food Science”: Professor Xie Jing et al. from Shanghai Ocean University: Research Progress on the Initial Adhesion Regulation Mechanism of Biofilms and Its Application in Food Quality Control
“Food Science”: Professor Hu Qiuhui et al. from Nanjing University of Finance and Economics: Effects of High-Temperature Storage Conditions on the Deterioration of Peanut Oil and Protein Quality
“Food Science”: Associate Researcher Hu Lihua et al. from Jiangsu Academy of Agricultural Sciences: Effects of Mildly Acidic Electrolyzed Water on Chlorophyll Degradation in Post-Harvest Broccoli
“Food Science”: Professor Qian Jing et al. from Jiangnan University: Effects of Gaseous Ozone Combined with Microperforated Packaging on the Quality and Antioxidant Properties of Waxberry
“Food Science”: Professor Wang Guangqiang et al. from Shanghai University of Technology: Research Progress on Lactic Acid Bacteria as Delivery Carriers for Bioactive Substances in Vivo
