Hello everyone, this week I would like to share a paper published by our research group in the Journal of the American Chemical Society, titled Photoactivatable RNA Tags for Subcellular Photolabeling of RNA. The research was conducted in collaboration with the Xu Pingyong and Zhang Jinyang research groups from the Chinese Academy of Sciences. Our group focuses on imaging RNA within cells, biosensors, and drug delivery.

Abstract
Although photactivatable fluorescent proteins have enabled precise analysis of subcellular protein dynamics, there is still a lack of similar photactivatable techniques for labeling subcellular RNA. To address this issue, we developed a photactivatable fluorescent RNA tag named PA-Broccoli, which mimics the function of photactivatable green fluorescent protein (PA-GFP). Similar to PA-GFP, PA-Broccoli is non-fluorescent in its initial state but emits strong green fluorescence upon exposure to ultraviolet light. Notably, PA-Broccoli exhibits superior performance: its fluorescence signal can be enhanced by 6000-fold after complete photactivation, and the activation speed is extremely fast (t₁/₂ ≈ 3 s), surpassing PA-GFP by one to two orders of magnitude in sensitivity and rate. Using PA-Broccoli, we made a series of important discoveries: first, we revealed that the mobility of cytoplasmic RNA is more restricted compared to proteins. Second, we found that circular RNA (circRNA) is exported from the nucleus via a Ran-GTP dependent pathway, and its accumulation rate in the cytoplasm is slower than that of linear mRNA. Finally, real-time imaging of mRNA within stress granules indicated that the exchange process of mRNA is energy-dependent.
Background
Since the discovery of green fluorescent protein (GFP), gene-encoded fluorescent tags have become core tools for monitoring the spatiotemporal dynamics of proteins. However, traditional fluorescent proteins have limitations due to constitutive fluorescence, making it difficult to track the dynamic processes of specific protein subpopulations. To overcome this limitation, photactivatable fluorescent proteins (PA-GFP) were developed. They can be selectively “turned on” by light, allowing for precise spatiotemporal tracking of processes such as protein transport and organelle interactions, revolutionizing research in related fields.
Similar to protein studies, the need for visualizing key intracellular molecules like RNA has also led to the development of various fluorescent RNA tags. These tags are typically RNA aptamers that can bind to non-fluorescent small molecules and activate their fluorescence, enabling imaging of RNA within live cells. Although these tools have advanced RNA functional studies, they face the same dilemma as traditional fluorescent proteins—constitutive fluorescence. This characteristic poses a significant challenge for the precise study of dynamic processes such as RNA localization and transport. Therefore, developing a photactivatable RNA tag to fill this critical technological gap has become an urgent need in the field of RNA biology.
To meet this demand, our study is based on the Broccoli RNA aptamer (a “RNA version of GFP” that can bind and activate GFP-like fluorophores), and we successfully designed and developed a photactivatable fluorescent RNA tag—PA-Broccoli—by chemically modifying the fluorophore with a light-sensitive cage. This tag mimics the optical switching characteristics of PA-GFP: it exhibits almost no fluorescence before ultraviolet activation and produces a strong signal afterward.
Remarkably, PA-Broccoli significantly outperforms its protein prototype PA-GFP in key performance metrics. Its fluorescence activation factor reaches approximately 6000-fold (compared to about 15-fold for PA-GFP), and its activation speed is much faster, with a half-life (t₁/₂) of approximately 3 seconds (compared to about 50 seconds for PA-GFP). In summary, PA-Broccoli is not only a successful optical switch but also represents a significant advancement in sensitivity and reaction kinetics over existing technologies.
Results
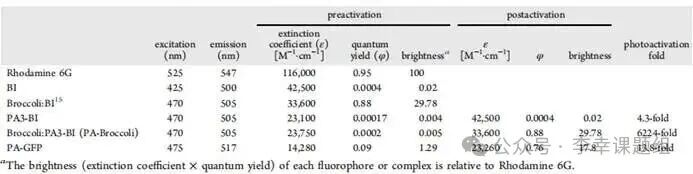
In this study, we selected the Broccoli RNA aptamer as the reporting motif. Broccoli can bind and activate the fluorescence of various GFP-like fluorophores. After comparing multiple candidate fluorophores (Table 1), we ultimately selected the BI molecule due to its optimal overall performance. The key advantages of the BI molecule include its extremely high signal-to-noise ratio: high brightness—when combined with Broccoli, its fluorescence quantum yield is very high, resulting in bright signals (φ=0.88). Low background: in its free state, its fluorescence signal is very weak (φ=0.0004), and there is minimal non-specific binding in cells. This significant difference in fluorescence signals before and after binding arises from the BI molecule’s similar “aggregation-induced emission” (AIE) characteristics—its intramolecular rotation is suppressed upon binding with Broccoli, resulting in strong fluorescence emission. Thanks to these excellent characteristics of low background and high brightness, the Broccoli:BI system was chosen as the ideal platform for constructing a new photactivatable RNA tag.
Table1 Photophysical and binding properties of different in vitro FLAPs
To convert the Broccoli:BI system into a photactivatable mode, the researchers proposed a design principle based on a “light-caging” strategy. The inspiration for this design came from the crystal structure of Broccoli RNA, which showed that the hydroxyl group of the BI fluorophore is located within a closed internal space of its binding pocket. Based on this, the research team speculated that if a large light-sensitive group (i.e., a “cage”) were attached to this hydroxyl group, the resulting steric hindrance would effectively prevent BI from binding to Broccoli, keeping the fluorescence in the “off” state. When the light cage group is removed by ultraviolet light, the BI molecule can restore its original structure, rebind to Broccoli, and activate fluorescence, thus turning the signal “on” (Figure 1A).
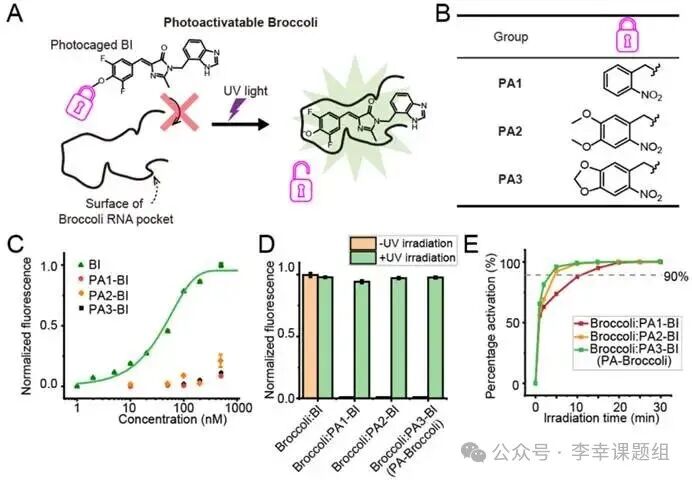
Figure1 Activation of fluorescence using Broccoli RNA tags
To validate the design principle, the researchers first connected three different light-cage groups to the hydroxyl group of the BI molecule, synthesizing PA1-BI, PA2-BI, and PA3-BI (Figure 1B). 1. “Off” state validation: The light-cage group effectively inhibits binding. Fluorescence titration experiments confirmed that prior to photactivation, the binding affinity of these three derivatives to Broccoli is extremely low. Their dissociation constants (KD) are over 1000 times higher than that of unmodified BI (KD=56 nM), indicating that the light-cage group successfully prevents their binding, keeping the fluorescence system effectively in the “off” state (Figure 1C). 2. “On” state validation: Achieving ultra-high fluorescence enhancement. After exposure to ultraviolet light, the fluorescence signals of all three systems were successfully activated, achieving approximately 6000-fold enhancement at the emission peak of 505 nm. This demonstrates that PA-BIs have extremely low background signals and very high brightness after activation, making them one of the highest contrast photactivatable fluorescent tags reported to date (Figure 1D). 3. Kinetic optimization: Screening for the fastest activating PA3-BI. To obtain the best performance tag, the authors further compared the photactivation rates of the three derivatives. The results showed that Broccoli:PA3-BI has the fastest activation speed, reaching about 90% of maximum fluorescence within 3 minutes. In contrast, PA2-BI and PA1-BI required 5 minutes and 10 minutes, respectively, to reach the same activation level (Figure 1E). HPLC-MS analysis also confirmed that PA3-BI removes the light-cage group the fastest. Based on its fastest activation rate, the authors ultimately selected the Broccoli:PA3-BI system for subsequent experiments and officially named it PA-Broccoli.
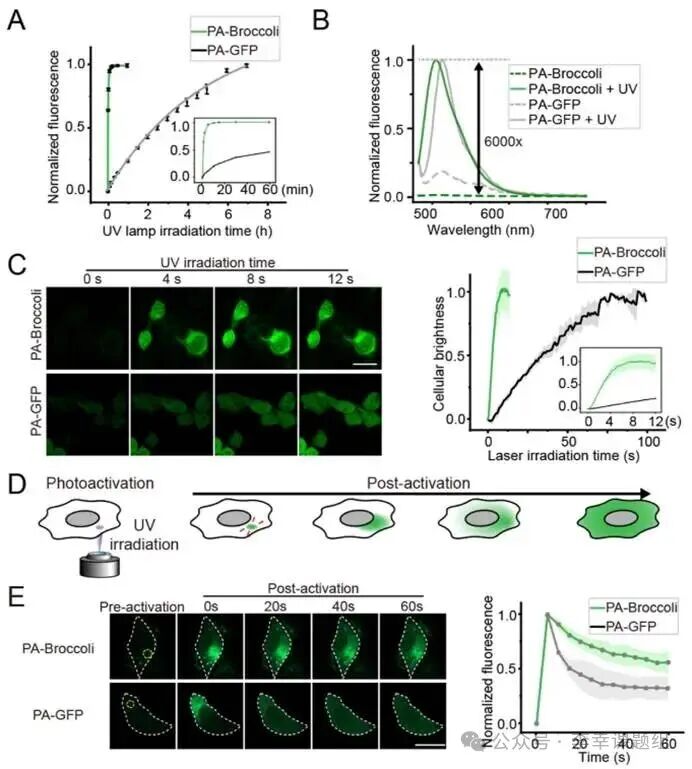
Figure2 PA-Broccoli exhibits superior high activation and rapid photactivation compared to PA-GFP as an RNA analog of PA-GFP.
In in vitro fluorescence assays, it was found that under the same ultraviolet light conditions, PA-Broccoli only requires3 minutes to reach 90% of maximum activation, while PA-GFP takes up to7 hours. In terms of fluorescence enhancement factor, PA-Broccoli achieves approximately 6000-fold, far exceeding PA-GFP’s approximately 13-fold (Figure 2A, 2B). In live cell imaging results, in HEK293T cells, the fluorescence activation half-life (t₁/₂) of PA-Broccoli is only about3 seconds, which is more than ten times faster than PA-GFP’s approximately 40 seconds, demonstrating its efficient and rapid response capability in real biological systems (Figure 2C).
In addition to its excellent activation efficiency, PA-Broccoli also possesses outstanding overall performance: superior photostability—thanks to its unique fluorophore “dynamic exchange” mechanism, PA-Broccoli is more resistant to photobleaching than PA-GFP, retaining about 60% of its fluorescence after 5 minutes of continuous exposure (PA-GFP retains only about 40%). Better biocompatibility: at effective activation doses, PA-Broccoli shows no significant phototoxicity and is suitable for both human and bacterial cells. Stronger versatility: this study confirms that this “light-caging” design strategy is also applicable to other RNA-fluorophore systems (such as PA-RhoBAST) and can even be extended to near-infrared (NIR) photactivatable systems with less biological damage, providing a feasible path for long-term, low-damage live cell RNA imaging.
PA-Broccoli’s powerful performance enables unprecedented high-precision spatiotemporal labeling of live cells. Whether for specific cell populations or for small regions of interest (ROI) within a single cell, the fluorescence signal can be precisely confined to the target area, with minimal diffusion or leakage over time (Figure 2D). In addition to high-precision spatiotemporal labeling, revealing restricted RNA movement, by locally activating at specific points in the cytoplasm, researchers found that PA-Broccoli-labeled RNA diffuses significantly slower than PA-GFP-labeled proteins (Figure 2E). Control experiments in fixed cells further confirmed that this phenomenon is due to the restricted movement of RNA molecules themselves, rather than dye diffusion. This tool exhibits extremely strong targeting capabilities, allowing for selective illumination of specific biomolecules. For example, researchers successfully achieved targeted labeling of U6 snRNA within Cajal bodies and were able to sequentially activate and track specific gene loci on different chromosomes within a single cell, fully demonstrating its powerful spatiotemporal specificity labeling capability.
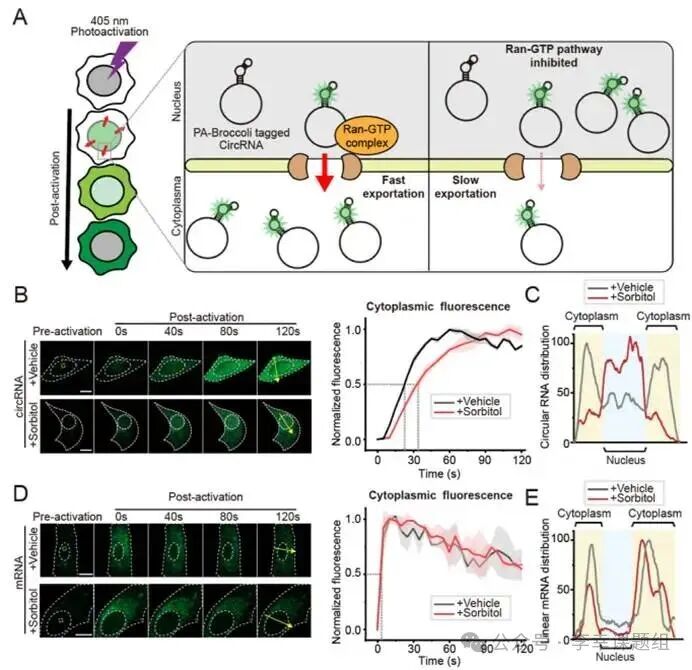
Figure3 Real-time imaging of PA-Broccoli reveals different nuclear export mechanisms of circRNA and mRNA
To demonstrate the application value of PA-Broccoli in elucidating complex biological processes, researchers used it to track the nuclear export dynamics of circular RNA (circRNA) in real-time. It is known that circRNA, after being synthesized in the nucleus, needs to be transported to the cytoplasm via the Ran-GTP pathway, but the specific dynamics of this process remain unclear. The research team activated only the molecules in the nucleus of U2OS cells expressing circular Broccoli RNA with a laser and then monitored the speed at which the fluorescence signal appeared in the cytoplasm to quantify its export rate (Figure 3A). In the experiment, the use of sorbitol to inhibit the Ran-GTP pathway significantly hindered the nuclear export of circRNA: its export half-life (t₁/₂) extended from about 22 seconds in the control group to about 32 seconds (Figure 3B). Meanwhile, fluorescence distribution maps showed that the inhibited circRNA was retained more in the nucleus (Figure 3C). These results intuitively demonstrate that the nuclear export of circRNA depends on the Ran-GTP pathway. In contrast, when the researchers conducted the same test on linear mRNA, they found completely different results: sorbitol treatment had no effect on the nuclear export rate and distribution of mRNA (Figure 3D, 3E). More importantly, the output speed of mRNA itself is much faster than that of circRNA, with a half-life of only about 3.7 seconds. In summary, the precise spatiotemporal labeling capability of PA-Broccoli successfully revealed that circRNA and linear mRNA not only rely on different nuclear export pathways but also exhibit significant differences in dynamics.
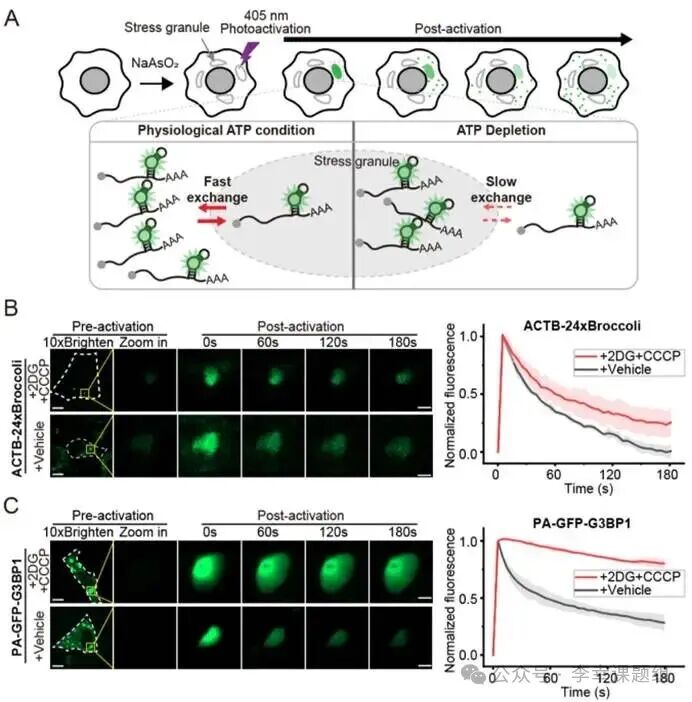 Figure4 mRNA within stress granules exhibits energy-dependent dynamics
Figure4 mRNA within stress granules exhibits energy-dependent dynamics
To explore the exchange dynamics of mRNA within stress granules (SGs), researchers utilized PA-Broccoli to observe this dynamic process. They first induced the formation of SGs in U2OS cells expressing Broccoli-labeled ACTB mRNA using sodium arsenite (NaAsO₂) (Figure 4A). The core of the experimental design was to compare the exchange rates of mRNA within SGs under normal and ATP-depleted conditions. In the ATP-depleted group, cells were treated with glycolysis inhibitors (2-deoxyglucose, 2DG) and oxidative phosphorylation uncouplers (carbonyl cyanide m-chlorophenyl hydrazone, CCCP). Subsequently, the researchers locally activated mRNA within individual SGs with a laser and recorded the fluorescence decay rate of that spot, which directly reflects the exchange speed of the labeled mRNA with the surrounding unlabeled mRNA (Figure 4A). Live cell imaging results showed significant differences in mRNA exchange rates under the two conditions: in the control group with normal ATP, the fluorescence signal from the photactivated mRNA rapidly decayed, with a half-life (t₁/₂) of about 26 seconds, indicating rapid exchange of mRNA in and out of SGs (Figure 4B). In the ATP-depleted experimental group, the fluorescence decay rate significantly slowed, with a half-life extending to about 60 seconds, demonstrating that mRNA exchange was inhibited (Figure 4B). To ensure that the observed phenomenon was indeed due to RNA exchange and not dye diffusion, a key control experiment showed that ATP depletion did not affect the diffusion rate of free BI dye within SGs. This strongly confirms that the exchange of mRNA within SGs is an active, ATP-dependent process. Finally, parallel experiments using PA-GFP on the SG core protein G3BP1 also observed a similar phenomenon: after ATP depletion, G3BP1 protein was also “locked” within SGs, with its exit rate from SGs significantly reduced (t₁/₂ extended from about 52 seconds to over 180 seconds) (Figure 4C). In summary, these results collectively demonstrate that ATP is crucial for maintaining the dynamic exchange of mRNA and proteins within stress granules.
Conclusion
This study successfully developed a high-performance photactivatable fluorescent RNA tag—PA-Broccoli. In key performance metrics, it comprehensively outperforms the classic PA-GFP, especially excelling in fluorescence enhancement factor (approximately 6000-fold), activation speed (t₁/₂ ≈ 3 seconds), and photostability. Utilizing this powerful tool, researchers made a series of important biological discoveries, including revealing that RNA exhibits a more restricted movement pattern in the cytoplasm compared to proteins. It clarified that circular RNA (circRNA) has a Ran-GTP dependent nuclear export pathway that is different from linear mRNA and slower in rate. It demonstrated that the dynamic exchange of mRNA within stress granules is an active process dependent on cellular energy (ATP). In summary, the emergence of PA-Broccoli fills a critical technological gap in the real-time tracking of RNA dynamic processes. It not only provides unprecedented opportunities for in-depth exploration of RNA localization, transport, and regulatory mechanisms at the subcellular level but also opens new research directions for understanding related life processes and disease mechanisms.
Authors:LHX
Reviewed by: SHZ ZXC
Original link:https://pubs.acs.org/doi/10.1021/jacs.5c07380