
▲ Click the image for details
PREFACE
Introduction
For Antibody-Drug Conjugates (ADCs), the selection of drug targets not only requires overexpression in tumors but also depends significantly on the efficiency of endocytosis, which is essential for drug release activity. In fact, the efficacy of ADCs relies on the efficiency of targeted-mediated internalization for delivering effective payloads into tumor cells.
The pathways and efficiency of ADC internalization are closely related to the efficacy and design of ADC drugs. This is because it is a crucial factor in determining whether the linker is cleavable, non-cleavable, or sensitive to pH/reduction, as well as whether the effective payload (or its active metabolites) can diffuse across the cell membrane to provide a “bystander effect,” and whether it enhances tumor killing rates or contributes to dose-limiting toxicity. Therefore, it is necessary to gain an in-depth understanding of the endocytic mechanisms of ADCs, as this is an extremely important first step for ADCs to exert their pharmacological effects in vivo.
01
Endocytic Pathways Related to ADCs

Generally, normal endocytosis can be divided into three stages: (1) budding formation, (2) membrane invagination and vesicle maturation, and (3) membrane fission and release into the cytoplasm. Various endocytic pathways have overlapping aspects, making the general process of endocytosis highly flexible and complex.
Clathrin-Mediated Endocytosis
Clathrin-mediated endocytosis (CME) is conceptually a simple process that involves several consecutive and partially overlapping steps. CME can be initiated structurally by certain receptors on the plasma membrane or requires ligand and/or antibody binding to initiate. CME begins when clathrin coat proteins start to accumulate on the inner leaflet of the plasma membrane. The coat proteins continue to assemble and grow by recruiting additional protein adapters from the cytoplasm and interacting with them. Key adaptor proteins facilitate membrane bending, concentrating the internalized receptor/ligand into a “clathrin-coated pit” (CCP). As the CCP invaginates, it separates from the plasma membrane through a fission process as the neck of the CCP narrows. Actin polymerization helps pull the CCP inward into the cytoplasm until fission is complete, releasing the CCP as a clathrin-coated vesicle (CCV). Finally, the CCV coat is disassembled, and the CCV fuses with endosomes to transport to specific subcellular locations or can be recycled back to the cell surface.
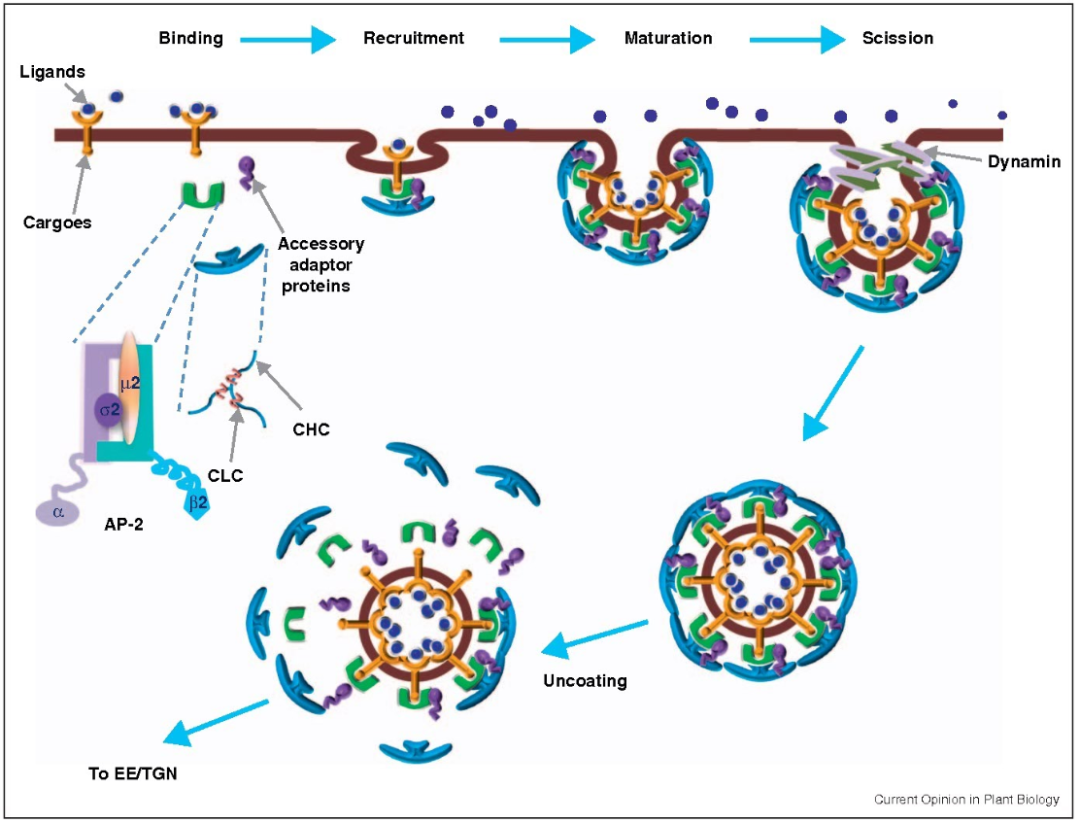
Clathrin is a key component of CME, composed of heavy and light chains. Three clathrin heavy and light chains form a trimer that interacts with other trimers to form a polygonal lattice around the emerging CCP. Adaptor protein 2 (AP-2) is a heterotetrameric complex that mediates the constriction of the CCP neck. Dynamin is a GTPase that forms helical polymers at the neck of the mature vesicle. After GTP hydrolysis, dynamin induces the fission of the vesicle from the plasma membrane.
Caveolae-Mediated Endocytosis
Endocytosis that does not rely on clathrin includes caveolae-mediated endocytosis, clathrin-independent carrier/GPI-enriched early endosomal compartments (CLIC/GEEC), and macropinocytosis.
Caveolae are flask-shaped invaginations of the plasma membrane characterized by high levels of cholesterol and sphingolipids, mediating endocytosis through a clathrin-independent pathway and are present in most cell types. The main scaffold protein of caveolae is caveolin, a complete membrane protein that forms oligomers of 20–24 kDa. Caveolin shares common scaffold domains that mediate interactions with itself and other proteins containing caveolin-binding domains.
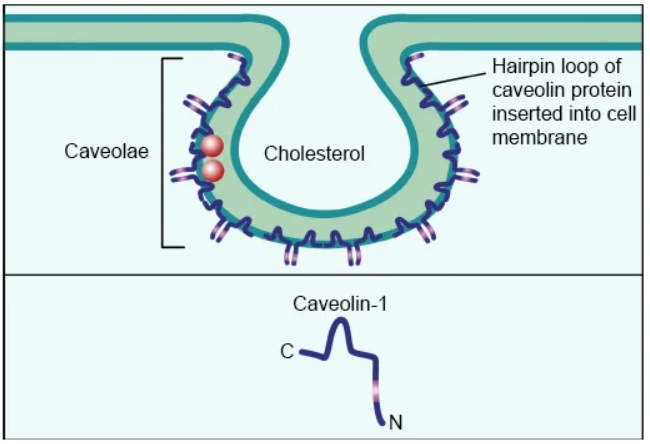
Although caveolae have a similar invagination morphology to CCPs, they are distinct. Simply put, the density of CCPs is constant, while the density of caveolae can vary significantly depending on the cell type. CCPs increase in size as budding endosomes mature, whereas caveolae vesicles maintain a constant size. Once inside the cell, caveolae form higher-order structures, rather than simple spherical endosomes formed by CCPs.
Another unique aspect of caveolin-mediated endocytosis is that only about 1% of caveolae bud from the plasma membrane. In a small fraction of internalized caveolae, it appears to follow a recycling pathway co-localizing with Rab5 (a marker for early endosomes). This may pose challenges for ADCs targeting receptors that utilize caveolin-mediated endocytosis.
CLIC/GEEC Endocytosis
CLIC/GEEC is an endocytic compartment that primarily occurs in ligand-activated cells, which may be triggered by growth factors, receptor cross-linking of antibodies, or bacterial toxins and viruses. Additionally, the cell membrane must be in a highly fluid state, as CLIC/GEEC does not function at sub-physiological temperatures or when the membrane is under higher tension.
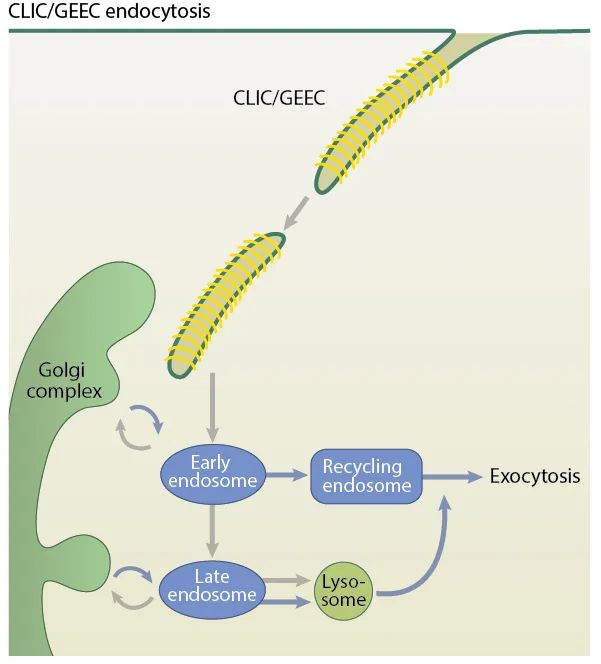
CLIC increases at the leading edge of migrating cells. Other relevant parameters for recognizing the CLIC/GEEC pathway include dynamin-independent membrane fission, sensitivity to cholesterol depletion, acquisition of Rab5/early endosome fusion, placental alkaline phosphatase (PLAP), and GTPase regulator associated with FAK (GRAF1).
Macropinocytosis
Macropinocytosis is a larger-scale form of endocytosis that typically involves highly folded regions/protrusions of the plasma membrane, which subsequently fuse with each other or with the plasma membrane. Membrane ruffling is a morphological feature of macropinocytosis.
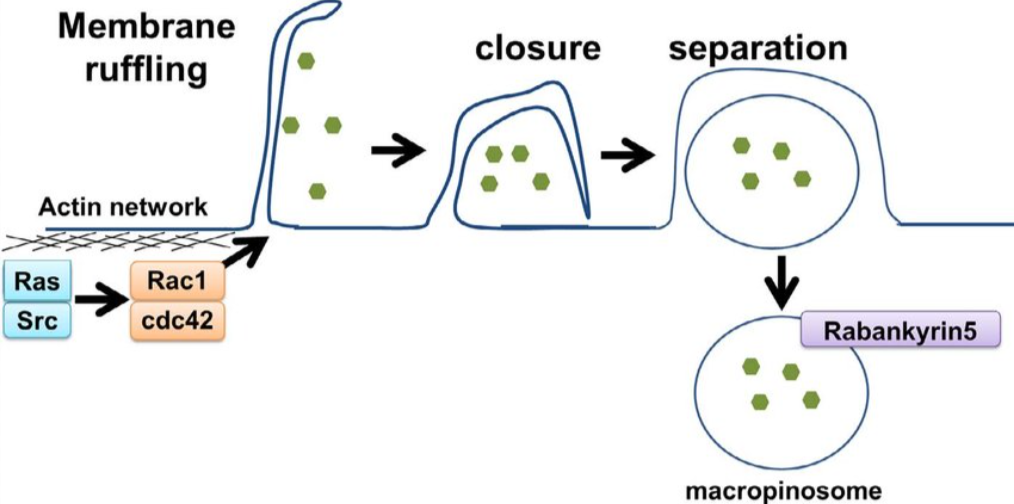
Macropinocytosis relies on actin polymerization, Rac1 protein, and p21-activated kinase 1 (PAK1). PAK1 is a key regulatory factor as it interacts with Rac1, which activates phosphatidylinositol-3-kinase (PI3K), Ras, Src, and Hsp90 to promote macropinocytosis. Macropinocytosis is also cholesterol-dependent, which is necessary for recruiting Rac1. These components ultimately lead to endocytosis with a larger uptake area than CME and caveolin-mediated endocytosis.
02
Endocytic Characteristics of ADC Target Antigens
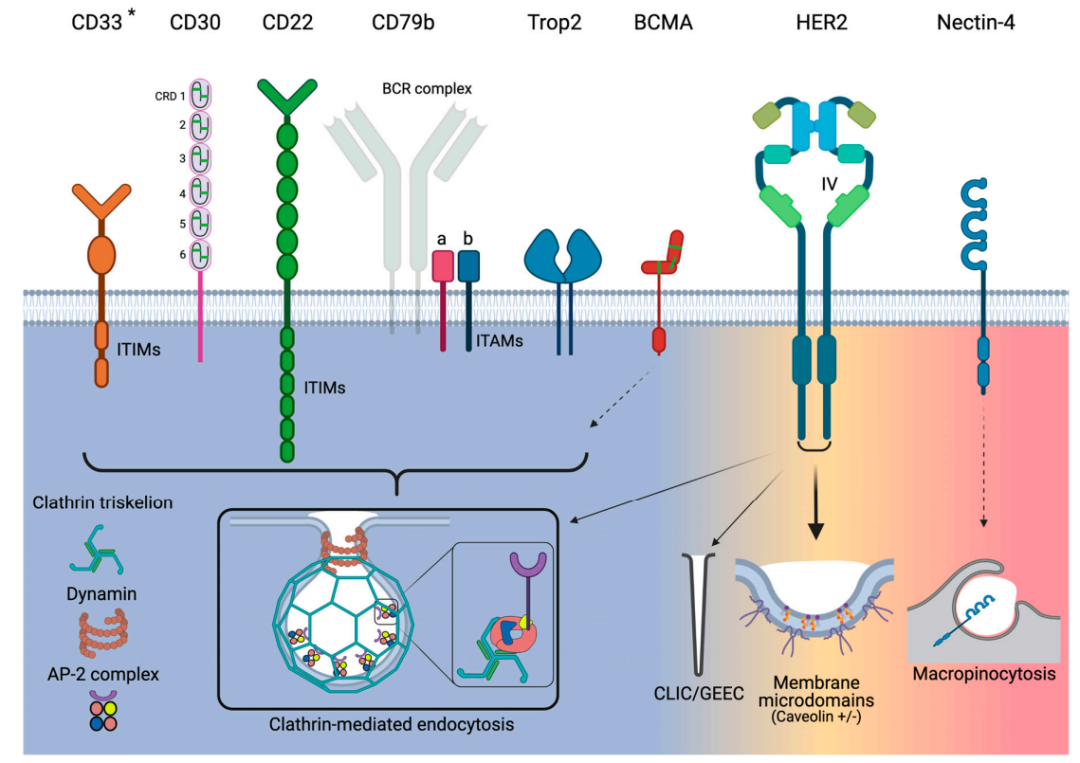
CD33
CD33 is a 67 kDa transmembrane glycoprotein receptor typically expressed on normal myeloid cells and is a target for GO due to its preferential overexpression on AML cells. The intracellular immune receptor tyrosine-based inhibitory motif (ITIM) of CD33 regulates its endocytosis, which can be activated through CME. There is no correlation between the expression level of CD33 in AML cells and its endocytic rate. CD33 is a slowly internalizing antigen, and additionally, cross-linking CD33 does not improve its endocytosis. Non-responsive AML patients to GO may be related to dysfunctional endocytosis of the CD33 receptor.
CD30
CD30 is a 120 kDa transmembrane glycoprotein belonging to the tumor necrosis factor receptor (TNFR) superfamily. Its extracellular portion consists of six extended conformation cysteine-rich domains (CRDs). CD30 is expressed on activated T and B cells as well as various lymphomas (including Hodgkin lymphoma and ALCL).
CD30 does not undergo endocytosis; instead, it is shed due to proteolytic cleavage mediated by matrix metalloproteinases (MMPs). Shedding is a characteristic of CD30 biology, and high concentrations of circulating soluble CD30 can serve as a serum marker for monitoring tumor progression. For the efficacy of ADCs, elevated circulating levels of CD30 seem to sequester injected ADCs, thereby reducing the number of ADCs that can localize to CD30-positive tumor sites. Therefore, the lack of endocytosis indicates that CD30 is not an ideal ADC target.
CD22
CD22 is a 140 kDa transmembrane glycoprotein, and like CD33, it is a member of the Siglec family and shares several structural features with that family. The key difference is that CD22 is much larger than CD33, as it has multiple Ig domains and ITIM/ITIM-like motifs. CD22 expression is restricted to B cells, and its expression level is elevated in most progenitor cells of various B cell malignancies (including ALL).
CD22 undergoes endocytosis via CME. Natural ligands rapidly internalize through structural interactions with CD22. These ligands are classified and degraded in lysosomes, while CD22 recycles back to the cell surface. Furthermore, CD22 ligand-induced endocytosis activates intracellular pools, replenishing or increasing the expression levels of CD22 on the cell surface. Therefore, CD22 has good endocytic characteristics for ADCs.
CD79b
CD79b is expressed only in immature and mature B cells and is overexpressed in ≥80% of malignant B cells. CD79a and CD79b are two non-covalently associated transmembrane proteins that mediate signaling and endocytosis. For the latter, the CD79a-CD79b heterodimer is the scaffold controlling BCR endocytosis. BCR endocytosis is primarily accomplished by CME and mediated by AP-2. Interestingly, CD79a directly interacts with the μ subunit of AP-2, thereby activating CD79b and leading to the endocytosis of the entire BCR complex.
Additionally, for ADCs, CD79a can be internalized as a monomer, but CD79b cannot. If the proximal membrane tyrosine (Y195) of CD79b is mutated, the binding of AP-2 to CD79a is blocked, and endocytosis is also blocked. In 18% of activated B cell-like DLBCL specimens, Y195 is mutated. In summary, there is evidence that the endocytic activity of CD79b depends on the internalization of the entire BCR complex rather than as a monomer.
TROP-2
Trop2 is a 46 kDa monomeric glycoprotein with selective overexpression, structural endocytosis, and lysosomal targeting characteristics, making it a very attractive target for ADCs. The internalization mechanism of Trop2 is related to CME.
The observed robust endocytosis of Trop2 may potentially be explained by significant Trop2 clustering. Studies on the conformational dynamics of Trop2 have found that Trop2 forms natural homodimers through interaction segments composed of the amino acids “VVVVV” located in the transmembrane domain. The dimerization of Trop2 can further recruit other cell surface proteins to bring Trop2 monomers closer together. Therefore, Trop2 clusters are likely formed by multiple dimers connected through lipid rafts and other membrane-associated proteins.
Trop2 binds to various ligands such as claudin-1, claudin-7, cyclin D1, and IGF1; however, none of these ligands have been shown to be internalized when binding or interacting with Trop2. Thus, the endocytosis of Trop2 is more pronounced in tumor cells compared to normal cells, indicating that Trop2 is a good target for ADCs.
BCMA
BCMA or CD269, also known as TNFR superfamily member 17, transmits signals that induce B cell survival and proliferation. BCMA has a molecular weight of only 20.2 kDa, and its ligand-binding extracellular region has a “chair-like” conformation composed of six CRDs. In addition to multiple myeloma, BCMA is also expressed in many hematologic malignancies, such as Hodgkin lymphoma and non-Hodgkin lymphoma.
However, little is known about the precise endocytic pathways utilized by BCMA. Regarding endocytosis, sialylation is a regulatory function that may induce BCMA to utilize CME for endocytosis.
HER2
HER2 is a 185 kDa transmembrane glycoprotein belonging to the EGFR family. Amplification of the HER2/neu gene is a known driver of human malignancies and metastasis. Due to the role of HER2 in cancer, it has been targeted for therapy for decades. HER2 has also been a target for ADCs, with T-DM1 and T-DXT approved for HER2-positive metastatic breast cancer patients.
There are various mechanisms for HER2 endocytosis, primarily CME, where co-immunoprecipitation clearly shows that HER2 directly binds to AP-2. Additionally, dynasore can completely block HER2 endocytosis in SKBR3 cells; secondly, caveolin-binding motifs φxφxxxxφ (φ represents aromatic amino acids Trp, Phe, or Tyr) are typically found in caveolin-associated proteins, interestingly, the sequence WSYGVTIW has been identified in the intracellular kinase domain of HER2; furthermore, studies have shown that HER2 can utilize the CLIC/GEEC endocytic pathway.
These different findings reveal important characteristics of HER2 endocytosis. Firstly, the endocytosis of HER2 is mixed; secondly, the caveolin-mediated endocytic pathway seems to be more frequently utilized.
Nectin-4
Nectin-4 is a 66 kDa type I transmembrane protein primarily involved in promoting cell-cell contact. Nectin-4 is an attractive target for ADCs as studies have shown that it is overexpressed in several tumor types but is almost absent in normal adult tissues.
Currently, no information has been found regarding the endocytosis of natural ligands or mAb/ADC complexes with nectin-4; however, studies on the endocytosis of nectin-4 binding pathogens can provide insights. Nectin-4 is also a receptor for the measles virus, and studies have shown that the measles virus enters MCF7, HTB-20 breast cancer, and DLD-1 colorectal cancer cells via macropinocytosis. Viral entry requires PAK1, whereas the dynamin inhibitor dynasore does not affect viral entry. Furthermore, cells expressing dominant-negative caveolin do not eliminate viral endocytosis.
Based on these indirect studies, nectin-4 exhibits strong endocytic activity required for viral receptors.

Note: The information is reproduced from various company websites and public information sources.The copyright belongs to the original author and is for readers’ learning, research, or appreciation only. It may not be used for commercial purposes. If there are issues regarding the content, copyright, or other matters, please contact for deletion within 30 days;This public account maintains neutrality regarding the content, statements, and opinions of the reproduced and shared materials and does not provide any express or implied guarantees regarding the authenticity or completeness of the content. This article is for industry analysis only and does not constitute investment advice. The stock market has risks, and investment should be cautious.
Consult BD for trading cooperation and brand promotion
Please add the assistant
(Please indicate your name + company )
)


 ▲ Click the image link for details
▲ Click the image link for details
END
↓ Click the QR code to follow the public account

Tap “Looking”
Encourage yourself on the path of growth
Take a step forward