
Although the number of interneurons in the hippocampus accounts for only 10-15% of the total cell population, their γ-aminobutyric acid (GABA) inhibitory circuits play an important role in the memory functions of the hippocampus.
In recent years, as more and more studies focus on the dynamic organization of inhibitory circuits in the hippocampus and their roles in hippocampus-dependent memory and learning, it is essential to understand the types of interneurons in the hippocampus and their influence on hippocampal function and neural plasticity. Professors Lisa T and Suhel T summarized these issues and published their article “The role of inhibitory circuits in hippocampal memory processing” in Nature Reviews Neuroscience in May 2022.


Welcome to join the
NationalLearning and MemoryAcademic Discussion Group
Add the editor’s WeChat
brainnews_11
-Message: Learning and Memory Research Group-

Hippocampal Inhibitory Interneurons
Hippocampal GABAergic interneurons exhibit high heterogeneity, which can be divided into four main categories based on their connectivity patterns and target cell locations: interneurons targeting the cell bodies of pyramidal cells (PCs) (ST), interneurons targeting the axon initial segments or axon-axon sites of pyramidal cells (AAC), interneurons targeting the dendrites of pyramidal cells (DT), and specific interneurons selectively targeting GABAergic cells (IS).
ST and AACs control the firing timing and rhythm synchronization of pyramidal cells at different oscillation frequencies.The main components of ST are parvalbumin (PV) + basket cells (PV-BCs), which belong to fast-spiking cells (Figure 1.a,f; Table 1).
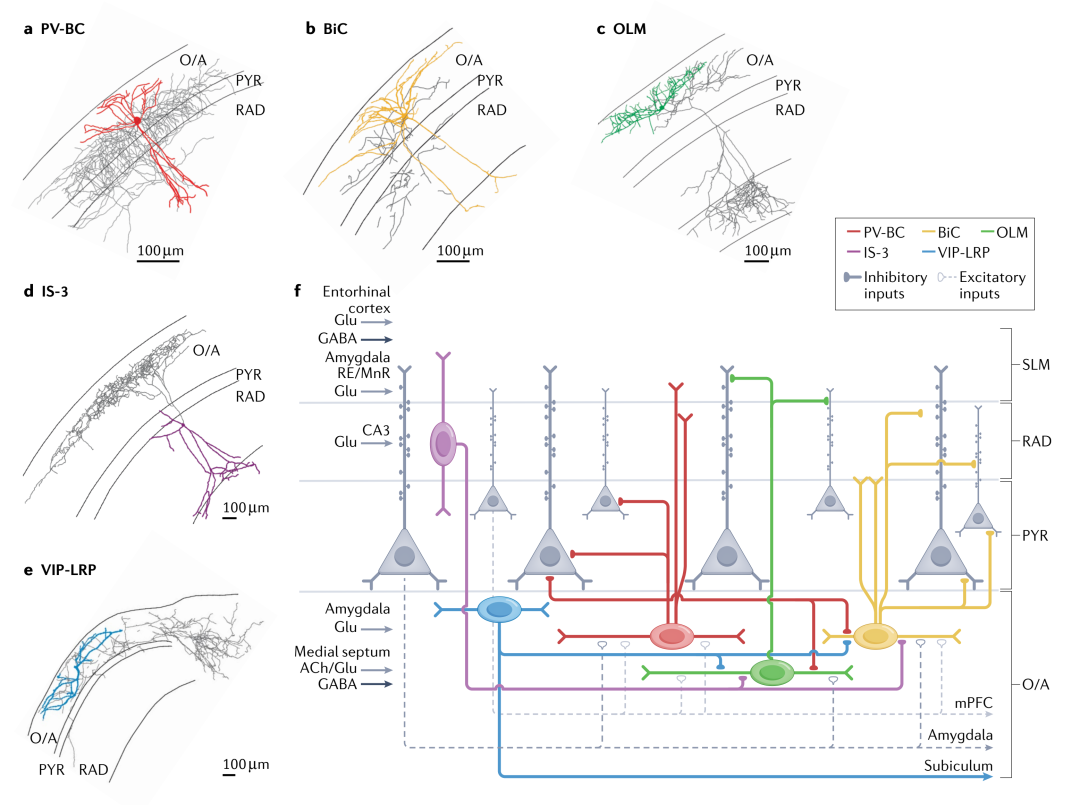
Figure 1. Main types of hippocampal CA1 inhibitory interneurons expressing PV, SOM, and VIP
PV-BCs receive a high density of excitatory inputs and provide different feedforward and feedback inhibition to PCs, with PV-BCs providing stronger inhibition to deep projecting PCs to the amygdala, but receiving stronger excitation from superficial PCs projecting to the medial prefrontal cortex (mPFC) (Figure 1.f).
DT sends synapses to the dendrites of PCs, controlling and integrating the excitatory input to PCs. DT includes many morphologically and molecularly diverse cell types, with a focus on the neuropeptide somatostatin (SOM) expressing DT cells, primarily BiCs and OLM cells.In addition to SOM, they also express PV and neuropeptide Y, these cells inhibit PCs and nearby interneurons’ proximal tips and basal dendrites, receiving inhibition from PV+ cells, and their physiological characteristics are similar to PV-BCs (Figure 1.b,f; Table 1).
OLM cells inhibit the distal dendrites of CA1 PCs, coordinating the excitatory input at the terminal in the stratum lacunosum-moleculare (SLM). They receive excitatory input from CA1 PCs, displaying dendritic spike and potential resonance characteristics (Figure 1.c,f; Table 1).
IS interneurons may gate information flow through different microcircuits, thus coordinating the activity of the entire network. In the CA1 region, IS are divided into four types, among which IS-3 cells express vasoactive intestinal peptide (VIP) and calbindin (CR), which send synapses to SOM+OLM cells and BiCs, representing the most excitatory cells in the hippocampus (Figure 1.d,f; Table 1). Additionally, long-range projecting IS (LRP-IS) express VIP and two types of muscarinic receptors, sending synapses to CA1 and the subiculum (Figure 1.e,f; Table 1).
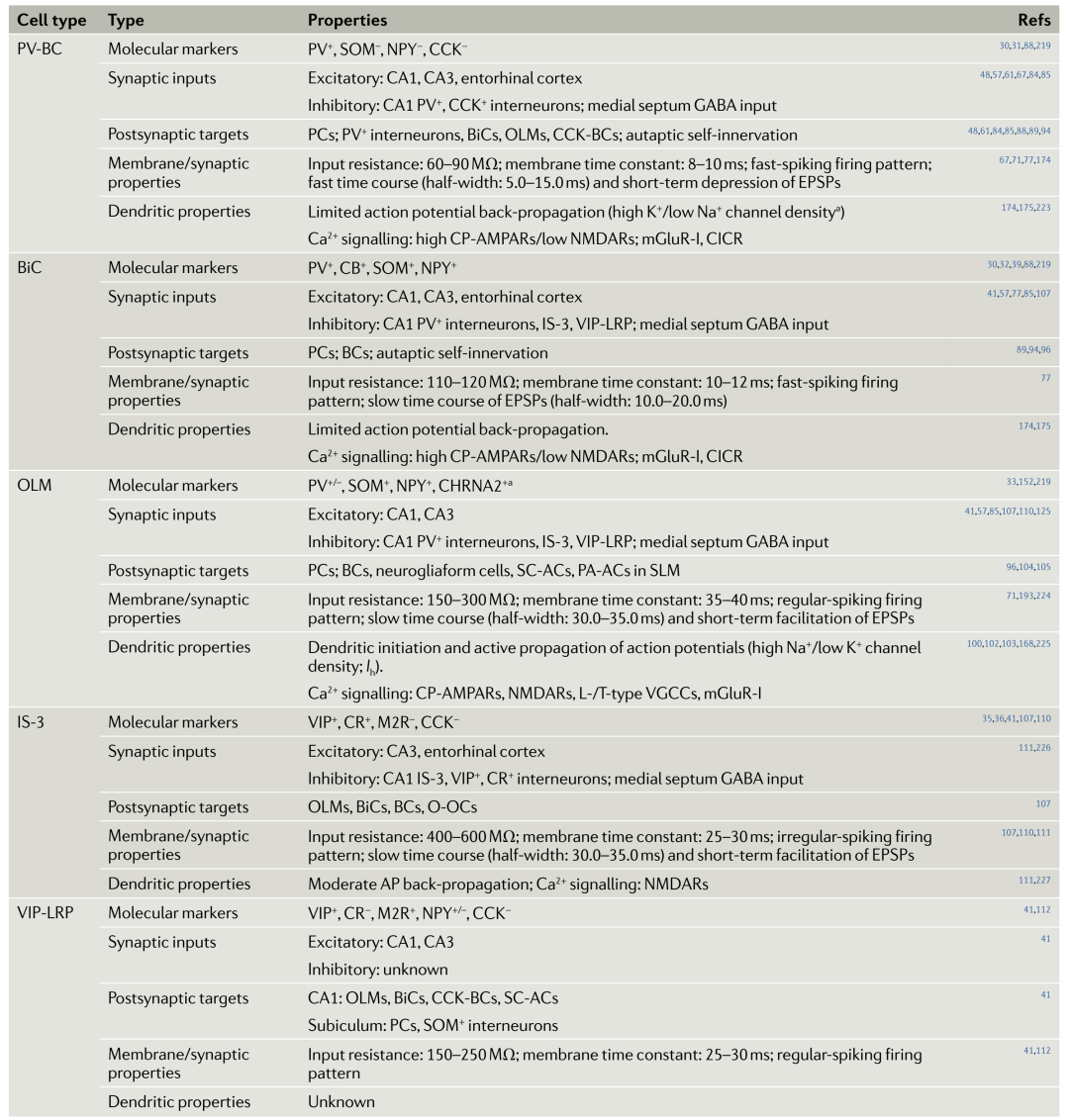
Table 1. Cell types and characteristics of CA1 PV+, SOM+, and VIP+ interneurons
Interneurons and Hippocampal Network Rhythms
There are rhythmic neural oscillations in the hippocampus caused by synchronized neurons, divided into θ oscillations (4-12Hz), γ oscillations (fast: 60-100Hz; slow: 25-55Hz), and sharp-wave ripples (SWRs, 110-200Hz). These oscillations are closely related to the encoding, consolidation, and retrieval of memories. Hippocampal network oscillations result from the continuous competition between excitation and inhibition, where inhibitory interneurons coordinate the excitation of pyramidal cells and synchronize their activity within specific time domains.
In recent years, with more refined control of neurons, it has been found that differentiated interneurons exhibit cell-type specificity and temporal specificity in regulating oscillations, for instance, PV-BCs are always active during the descending phase of θ oscillations and during SWRs, while IS-3 cells increase activity at the peaks of θ waves, and VIP-LRP remain silent during θ oscillations, with neither activating during SWRs.
Moreover, studies have found that PV+ interneurons are key factors in regulating hippocampal θ and γ rhythms, while the role of SOM+ cells remains controversial (Figure 2).
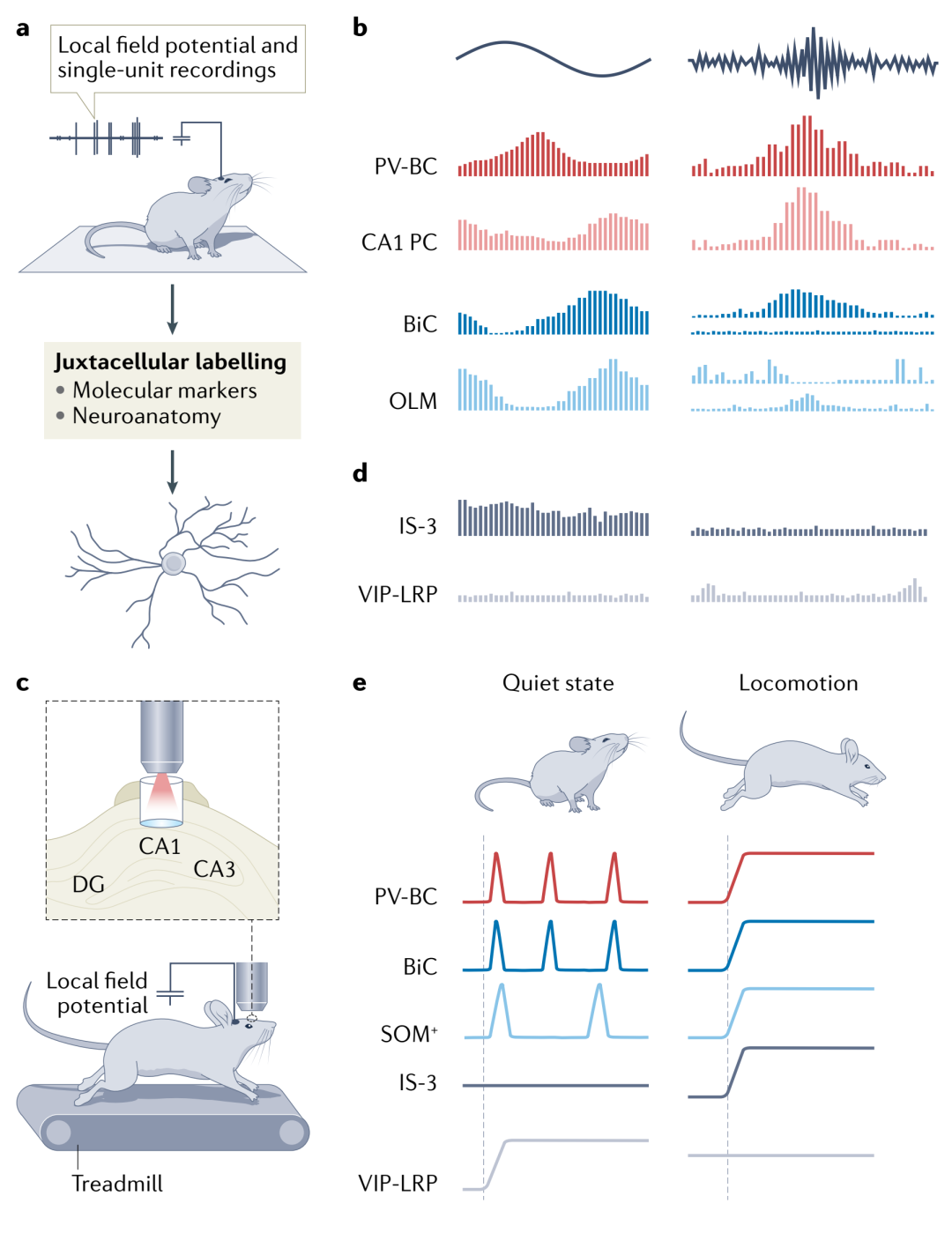
Figure 2. Activity patterns of CA1 inhibitory interneurons during hippocampal network oscillations
Synaptic Plasticity of Interneurons
Through different forms of synaptic plasticity, network oscillations generated by dynamic interactions between PCs and interneurons can lead to changes in network connectivity strength and firing output (Figure 3). For example, under θ oscillations, SOM+ cells can produce Hebbian long-term potentiation (LTP) (Figure 3.a), while ST PV+ cells produce LTP under γ oscillations (Figure 3.b). These changes depend on cell-type specific mechanisms.
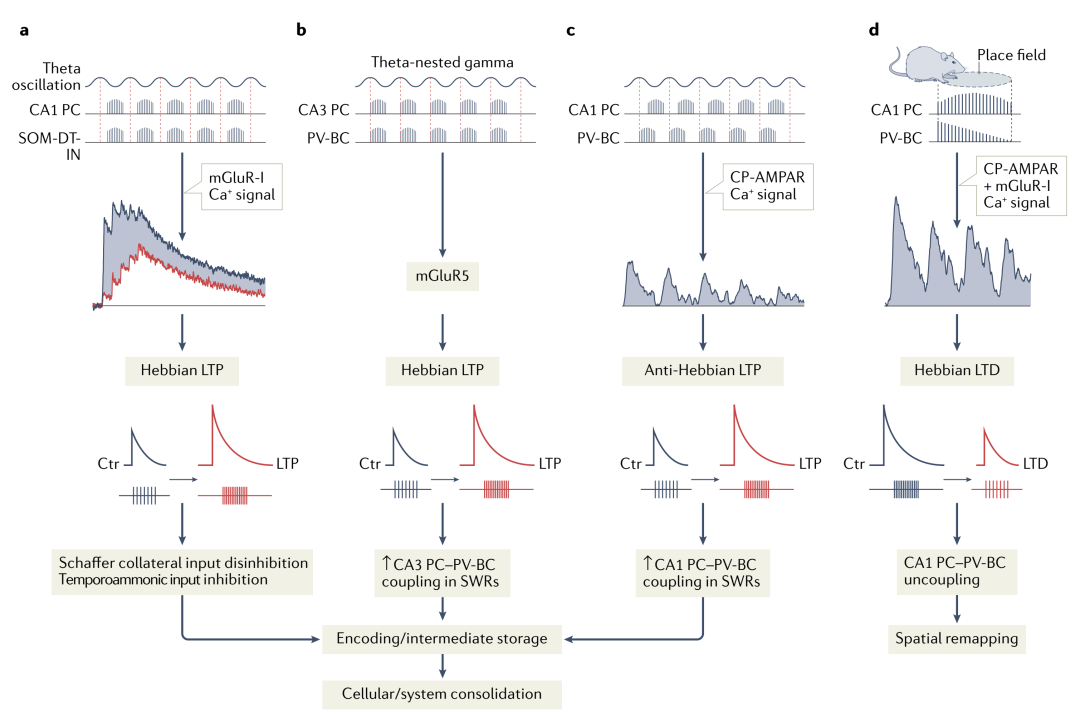
Figure 3. Activity-dependent synaptic plasticity of CA1 inhibitory interneurons
Role of Interneurons in Memory Encoding and Consolidation
Hippocampal PCs encode spatial information through firing frequency and timing relative to the θ phase. Increased firing rates at specific spatial locations lead some PCs to form place fields. CA1 PV+ cells, SOM+ cells, and VIP+ cells can modulate the firing rates and timing of PCs within place fields, thus regulating the encoding of spatial information. Interneurons also modulate the encoding of contextual memory (Figure 4), with contextual fear conditioning experiments (Figure 4.a) being a classic paradigm for studying mouse contextual memory.
The modified version of this experiment (head-fixation, accompanied by high-resolution effects and chemogenetic manipulation) found that silencing SOM+ cells inhibits fear learning, and SOM+ cells, especially OLM, can promote the encoding of contextual fear memory (CFM) by inhibiting negative stimulus-related entorhinal cortex inputs (Figure 4.c), while silencing PV+ cells inhibits the consolidation of CFM, specifically by inhibiting the coupling of hippocampal CA1 ripples (120-250Hz) with mPFC spindle waves (12-16Hz), thus inhibiting memory consolidation (Figure 4.d). Furthermore, inhibition of the mTORC1 pathway in SOM+ cells also leads to impaired memory consolidation.
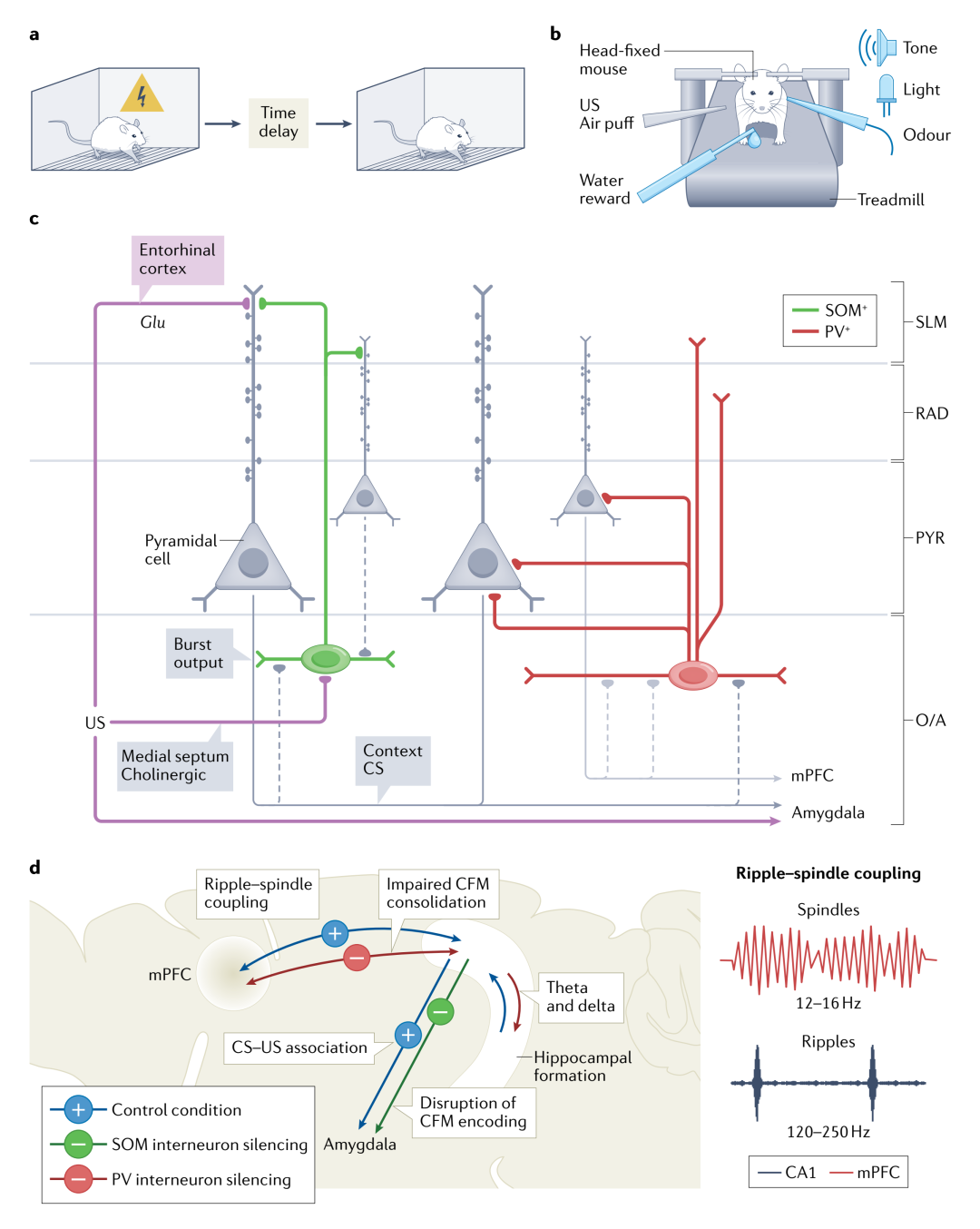
Figure 4. The role of hippocampal CA1 interneurons in contextual fear memory encoding and consolidation
Conclusion
Different types of hippocampal interneurons influence various aspects of memory formation by modulating the firing rates and timing of PCs, and identifying and understanding these neuron types and their underlying mechanisms is crucial for further research on hippocampal memory functions.
References
Lisa T and Suhel T. The role of inhibitory circuits in hippocampal memory processing. Nature Reviews Neuroscience, 2022. DOI: 10.1038/s41583-022-00599-0.
Compiled by: KK (brainnews creative team)
Reviewed by: Simon (brainnews editorial department)







