Song Wenjie1,2, Liang Yuzheng1,2, Tao Zhen1,2, Zhong Qingxiang1,2, He Yicong1,2
1.Sun Yat-sen University, School of Geography and Planning, Guangzhou, Guangdong 510006
2.Sun Yat-sen University, Field Observation Research Station of Carbon-Water Coupling in the Karst Forest Ecosystem of Northern Guangdong, Qingyuan, Guangdong 511500
1. Composition and Characteristics of Soil Organic Carbon (SOC) Pool
The SOC pool consists of plant-derived carbon and microbial-derived carbon, and there are differences in the composition and properties of different components.
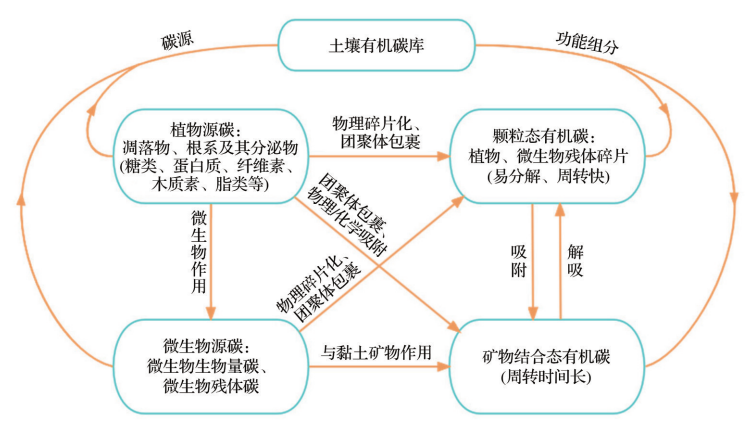
Composition and Properties of Soil Organic Carbon Pool
2. Microbial-Mediated Changes in SOC
Soil microbial activity is the main driving force for the formation, transformation, and stabilization of SOC, especially the microbial synthesis metabolic capacity and its activity rate directly affect the contribution of microorganisms to SOC.
(1)Microbial Transformation of Plant Carbon
The input of plant litter to the soil surface marks the transformation of plant carbon to soil carbon. This process is mediated by microorganisms, forming soil organic matter (SOM). According to Cotrufo’s dual pathway model, the formation of SOM mainly occurs through two pathways: one is that non-structural and low molecular weight dissolved organic matter (DOM) is absorbed by microorganisms, which after internal turnover combines with minerals to form stable organic carbon (MAOC); the other is that partially decomposed or undecomposed plant fragments form particulate organic matter (POM) through physical migration. Compounds in root litter are not easily decomposed and absorbed by microorganisms, mostly forming particulate organic carbon (POC) through external modification. In addition, microorganisms in the rhizosphere soil can synthesize metabolites that adsorb to minerals to form stable MAOC. However, when the SOM content in the soil is high, MAOC may reach saturation, while POC can continue to increase. When soil carbon is saturated, the dynamics of the stable SOC pool are mainly regulated by soil physicochemical protection mechanisms, while the continued burial effect of microorganisms can be neglected.
Moreover, lignin and cellulose are complex organic compounds found in the cell walls of vascular plants, characterized by high stability and resistance to degradation, making them relatively stable plant-derived carbon in soil. However, enzymes secreted by fungi can catalyze the oxidation of lignin, forming low molecular weight, highly soluble compounds. Some of these compounds are synthesized into microbial-derived carbon, while others are decomposed into CO2 and released from the soil carbon pool.
(2)Contribution of Microbial-Derived Carbon to SOC Pool
Soil microbial-derived carbon (MRC) is a major contributor to SOC, consisting of carbon content in microbial biomass (MBC) and residues and metabolic products after microbial death. MRC easily combines with minerals to form MAOC, making an important contribution to the stability of SOC.
Microorganisms have high substrate specificity, preferentially utilizing nutrient-rich substrates for synthetic metabolism. For example, Geobacter spp. tend to utilize simple carbon molecules like peptides and carbohydrates for glycolysis and the tricarboxylic acid cycle, with the energy and metabolic intermediates produced used for biosynthesis.
The soil microbial community mainly includes fungal and bacterial communities, producing MRC as fungal residual carbon (FRC) and bacterial residual carbon (BRC), respectively. On a global scale, the average content of soil MRC is 10.51 g/kg, making it a major contributor to SOC, accounting for 46.47% of SOC. In different ecosystems, the contribution rate of MRC to SOC varies, in the order of: grasslands (48.4%) > farmland (46.9%) > broadleaf forests (40%) > tropical rainforests (39%) > shrublands (32.27%) > coniferous forests (24.14%). This may relate to the properties of litter, climatic conditions, and microbial activity in different ecosystems.
In summary, the formation and contribution of soil MRC are influenced by various factors, including the metabolic characteristics of microorganisms, substrate specificity, types of ecosystems, and climatic conditions. Therefore, in-depth research on the mechanisms by which these factors affect the formation and contribution of soil MRC will help to better understand the cycling and stability mechanisms of soil organic carbon.
(3)Microbial-Mediated SOC Transformation
Stimulation and continued burial effects: The transformation of SOC by microorganisms involves both the degradation and mineralization processes that release CO2 and the assimilation and synthesis processes that form stable SOC. Thus, the dynamics of SOC mainly depend on the intensity of the two processes of microbial dissimilation and assimilation (MBC generation and MRC accumulation). The stimulation and continued burial effects in soil significantly impact the cycling and stability of SOC.
The stimulation effect refers to the process induced by the decomposition of litter and the input of root exudates in the soil, which can accelerate the decomposition of SOM, altering the size of the SOC pool, thus affecting global climate change. Among these, the rhizosphere stimulation effect is one of the main driving forces for SOM turnover and nutrient cycling, regulated by the availability of soil nutrients. Substances secreted by roots can stimulate microbial activity and enzyme secretion, thereby mineralizing SOC. Furthermore, the stimulation effect couples biological synergistic metabolism and mineral protective carbon transfer mechanisms.
However, despite the significant stimulation effect, due to the large biomass of fine root and the prolonged residence time of root litter in the soil, root carbon input can lead to a net increase in SOC. The stability of SOC formed from root carbon transformation is high because roots contain certain compounds that can suppress microbial colonization and invertebrate feeding, slowing down their decomposition process. Additionally, the direct contact of roots with soil minerals facilitates the rapid stabilization of their decomposition products.
On the other hand, the continued burial effect enhances the formation of MBC and the accumulation of MRC through the “internal turnover” pathway, continuously contributing stable organic carbon derived from microorganisms to the soil. This process promotes the stability and sequestration of SOC.
In conclusion, the balance between stimulation and continued burial effects regulates the storage and stability of the SOC pool.
The microbial community’s role in SOC transformation: The quantity and community composition of microorganisms are the main driving factors for the decomposition of litter. Different community compositions show significant differences in SOC metabolic processes and decomposition rates. Fungi secrete more extracellular enzymes that degrade SOM than bacteria; an increased ratio of fungi to bacteria significantly enhances SOM decomposition. Basidiomycetes can efficiently hydrolyze lignin, cellulose, and carbohydrates; Gram-negative bacteria degrade lignin faster than Gram-positive bacteria; ascomycetes are adept at decomposing carbohydrates without lignin degradation. Extracellular enzymes convert organic substrates into inorganic forms and large molecular substrates into soluble substrates for microbial assimilation, directly affecting the dynamic changes of SOC pools.
Compared to the mobility-limited bacterial community, the fungal community provides significant physical and chemical protection to SOC. The fungal community transfers litter-derived carbon to the soil through hyphal networks and preferentially accumulates in large aggregates contributing to POC; carbon exuded from fast-growing and high-yielding hyphae and hyphal residues significantly contributes to the MAOC pool; extracellular hyphae directly participate in the formation of soil aggregates. For instance, hyphae dying on the surface of conifer litter can co-precipitate with Mn-rich particles to form aggregates that enhance the physical protection of SOC. In forest soils dominated by arbuscular mycorrhizal fungi (AMF), the membranes formed by AMF covering the surface of mineral particles hinder water-mineral interactions, thus protecting MAOC. After AMF die, a glycoprotein secreted during turnover (glomalin-related soil protein) contributes to the formation and stabilization of soil aggregates, allowing SOC to be sequestered for the long term.
Additionally, mycorrhizal fungi can secrete specific enzymes to decompose nutrient-rich substrates to meet their growth nutritional needs, which leads to nutrient limitation for other decomposers in the soil, reducing their decomposition rates and increasing SOC storage; while ectomycorrhizal fungi (ECMF) utilize their extensive exogenous hyphal networks, allowing the carbon sequestration capacity of soil units in ecosystems dominated by ECMF-related plants to reach 1.7 times that of ecosystems dominated by AMF-related plants. ECMF has been shown to store SOC in the form of POC; while ecosystems dominated by AMF are more inclined to form MAOC due to rapid decomposition and MRC production.
3. Responses of Microbial-Mediated SOC Transformation to Environmental Changes
Environmental factors (temperature, humidity, texture, pH, etc.) respond to global changes, affecting microbial communities, metabolism, and enzyme activity, thereby regulating soil carbon cycling. Climate change impacts microbial properties and enzyme activity; for example, warming of the soil enhances ligninase and cellulase activity, altering the stability of inert carbon; warming of alpine meadows inhibits carbon-decomposing enzymes, favoring SOC sequestration.
Changes in soil moisture affect microbial respiration and metabolism; under high humidity, microorganisms reduce CUE and increase nitrogen use efficiency; dry conditions limit enzyme production and microbial biomass, favoring SOC retention.
Soil texture and mineral composition influence SOC retention and microbial distribution; clay particles form micro-aggregates on organic matter surfaces, with bacterial products interacting with minerals to form stable MAOC.
Soil pH complexly affects microbial community structure, with different microbial populations suitable for different pH ranges; pH and exchangeable aluminum regulate microbial energy flow and higher metabolic efficiency (CUE).
Global environmental changes lead to changes in the microbial regulatory mechanisms of SOC formation, transformation, and stability; annual precipitation, total nitrogen, and SOC changes affect microbial-derived carbon accumulation, subsequently altering SOC composition and properties.
Conclusion
Soil is the largest organic carbon pool in terrestrial ecosystems, with SOC reserves, components, and changes being complexly regulated by the coupling of plant carbon input, microbial activity, and soil matrix, where microbial properties play a key role in the process of plant carbon transforming into SOC. Currently, stable isotope labeling methods and biological labeling methods are widely used to trace the changes in SOC pool dynamics driven by microorganisms. Based on global environmental changes and ongoing related research, future studies on SOC pool dynamics should focus on:
In-depth research on the formation mechanisms of stable SOC functional components mediated by microorganisms at the plant-soil interface. The coupling of litter, microorganisms, and soil matrix drives the formation of SOM aggregates and MAOC. On the surface of litter, especially the surface of recently fallen litter, microbial activity is intense, and the products of microbial activity (extracellular polymers) drive the adhesion of fine-grained minerals to form stable MAOC and soil aggregates; these two processes and mechanisms for forming stable SOC are not affected by the mechanical composition of the soil. The soil-atmosphere CO2 exchange flux is constrained by the composition and properties of the surface SOC; therefore, the reserves, composition, and stability of surface soil SOC should be a focus of attention.
Under the influence of global environmental changes, the composition, metabolic levels, and functions of soil microbial communities constrained by soil environments and their compositional elements have also changed. To gain a deeper understanding of SOC reserves, composition, and dynamics, future research should strengthen the exploration of microbial CUE and its environmental dependencies; construct a coupling model of surface environment-soil microbial activity-SOC stability to provide theoretical support for predicting SOC responses to climate change and long-term sequestration of SOC.

The “Earth Science Progress” is a comprehensive and academic journal co-sponsored by the Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, and the Earth Science Department of the National Natural Science Foundation of China, with Academician Fu Bojie as the editor-in-chief. The journal has been rated as a high-quality scientific and technological journal in China by the Ministry of Science and Technology for many years, and in 2020, it was selected into the high-quality scientific and technological grading catalog T1 area in the field of Chinese geology, and is included in important domestic and international databases such as the “Chinese Core Journal Overview”, “Core Version of the Chinese Science Citation Database” (CSCD), Scopus, EBSCO, and American Chemical Abstracts (CA).The publication fields involve:Astronomy, surveying and mapping, geophysics (solid Earth geophysics, hydrological sciences, and space physics), atmospheric sciences (meteorology), geology, geochemistry, oceanography, and geography.