Clicktheblue text to follow us
Hello everyone, today we present an article published in July 2025 in the Journal of the American Chemical Society titled “Inhibitor-Dependent Tolerance of New Delhi Metallo-β-Lactamase Driven by Single Mutation-Induced Conformational Changes.”
β-lactam antibiotics are among the most widely used antimicrobial agents in clinical settings, but their effectiveness is severely challenged by β-lactamases. Among these, metallo-β-lactamases are particularly concerning due to their ability to hydrolyze nearly all β-lactam antibiotics, including carbapenems. The authors deciphered the evolutionary strategy of New Delhi metallo-β-lactamase (NDM) mutants through conformational dynamics studies, utilizing hydrogen-deuterium exchange mass spectrometry (HDX-MS) to analyze the conformational dynamics of NDM in different inhibitor binding states. They revealed the critical role of the α3–L8 region in the allosteric regulation of NDM, elucidating how the M154L mutation in this region enhances NDM’s resistance by modulating conformational dynamics, providing a theoretical basis for the design of novel inhibitors based on conformational dynamics.
 1. Analysis of the Structure and Dynamic Features of NDM-1
1. Analysis of the Structure and Dynamic Features of NDM-1
The crystal structure of NDM-1 displays typical αβ/βα characteristics (Figure 1), with two central anti-parallel β-sheets flanked by two pairs of α-helices. The active site comprises five ASLs (Figure 1e), where ASL2, ASL3, and ASL5 participate in zinc coordination, while ASL1 (L3) and ASL4 (L10) accommodate large substrates (Figures 1b, c). The critical roles of the loops ASL1 and ASL4, located on either side of the two zinc ions, have been widely recognized in previous structural studies. The ASL1 and ASL4 regions in the crystal structure show significant chain alignment deviations, indicating conformational heterogeneity of NDM-1 in its native state. The authors first conducted a comprehensive analysis of the conformational dynamics of wild-type NDM-1 and the mutant NDM-M154 using HDX-MS technology: they found that the C-terminal domain is more prone to hydrogen-deuterium exchange than the N-terminal, indicating greater flexibility. The active site loops ASL2 and ASL3 (involved in zinc coordination) exhibited lower deuterium uptake, suggesting a more stable structure, while ASL4 and ASL5 (responsible for substrate ingress and egress) showed higher dynamics. A comparison with the B-factors from the crystal structure revealed a high consistency with the HDX-MS results, validating the reliability of this technique.
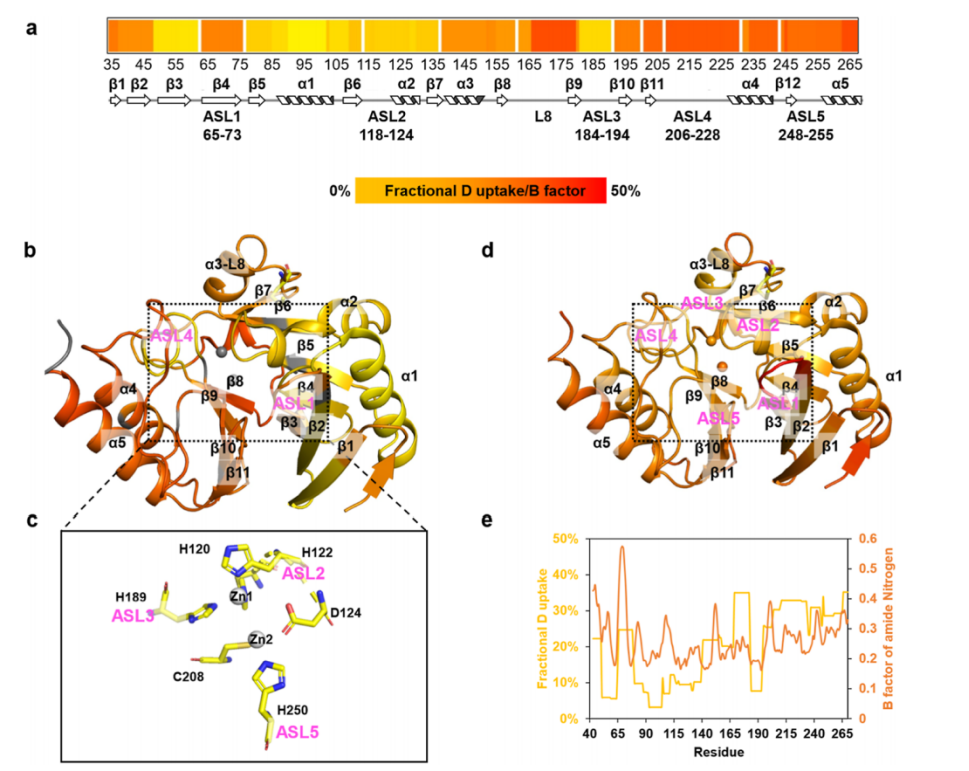
Figure 1
1) Zinc Intercalators: L- and D-Captopril Next, the authors studied the conformational changes of NDM induced by different inhibitors, starting with the zinc intercalator, captopril, which can non-covalently intercalate with the active site zinc ion to inhibit NDM-1, but these two drugs have significantly different IC50 values of 157.4 and 20.1 μM, respectively. The authors used HDX-MS to analyze the conformational dynamics as a potential determining factor for distinguishing the structural differences in the mechanisms of action between these stereoisomers. They found that after inhibition by l- and d-captopril, the HDX of NDM-1 underwent significant changes (Figure 2), where F67 (ASL1) and N220 (ASL4) formed hydrophobic and hydrogen bond interactions with captopril, indicating that inhibitor binding stabilizes these regions structurally. In contrast, residues 118–129 and 248–257 spanning ASL2 and ASL5, respectively, showed significantly less protected conformations (increased deuterium uptake), indicating that zinc intercalation leads to relaxation of the zinc coordination network. Previous crystal structures of NDM-1 in complex with L-captopril indicated that L-captopril’s thiol binds between the active site zinc ions, and the inhibitor does not directly contact the zinc-coordinating residues. The authors inferred that L-captopril could displace zinc coordination and energetically relax the constraints on ASL.
Surprisingly, the α3-L8 and C-terminal regions, which do not directly participate in inhibitor binding, exhibited significantly higher deuterium uptake, demonstrating more flexible conformations, suggesting that these regions become more flexible after inhibitor binding and may play a role in allosteric regulation. Previous work has rarely focused on the structural changes in the α3-L8 region located in the middle of the enzyme. It is speculated that due to the flexibility of this region, other structural methods may find it difficult to consolidate conformational changes. The authors’ HDX results indicate that the α3-L8 region may be crucial for the conformational flexibility of NDM-1.
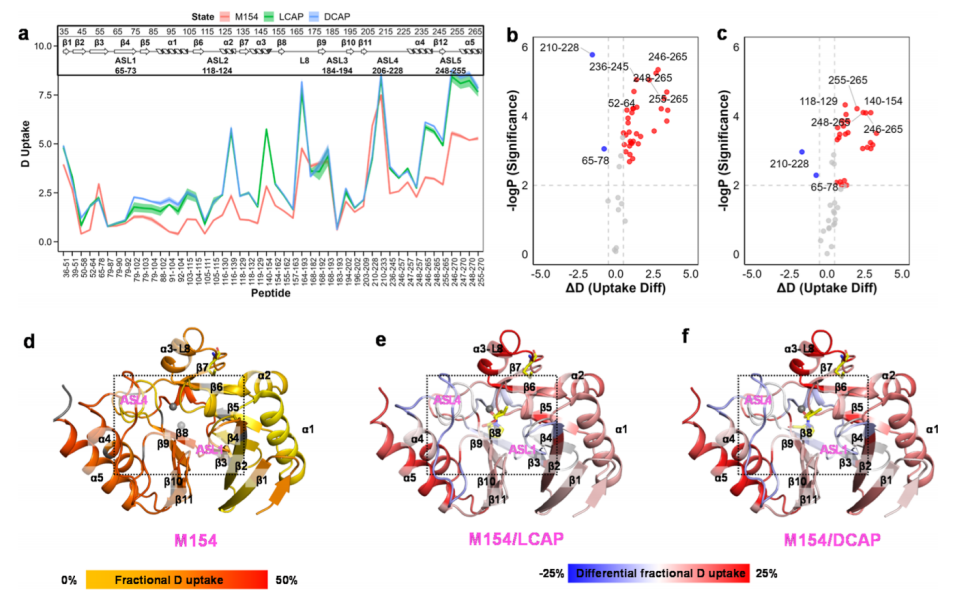
Figure 2
2) Zinc Chelators: AMA and Ebselen
AMA can chelate Zn2+ ions but cannot chelate Zn1+ ions, making it an effective inhibitor of NDM-1. Interestingly, the addition of excess zinc ions can restore activity in the presence of AMA. Ebselen is a covalent inhibitor that modifies C208 and strips Zn2+ after binding. The authors employed size-exclusion chromatography combined with native mass spectrometry (nSEC-MS) to analyze the structural changes of AMA and ebselen with NDM mutants (Figure 3). They found that both AMA and ebselen induced protective changes in the ASL3 and L8–β9 regions, indicating that the absence of Zn²⁺ actually enhances the stability of these regions. The authors observed that the α3–L8 region exhibited similar dynamic change patterns under all inhibitor actions, suggesting it may be a conserved allosteric regulatory region.
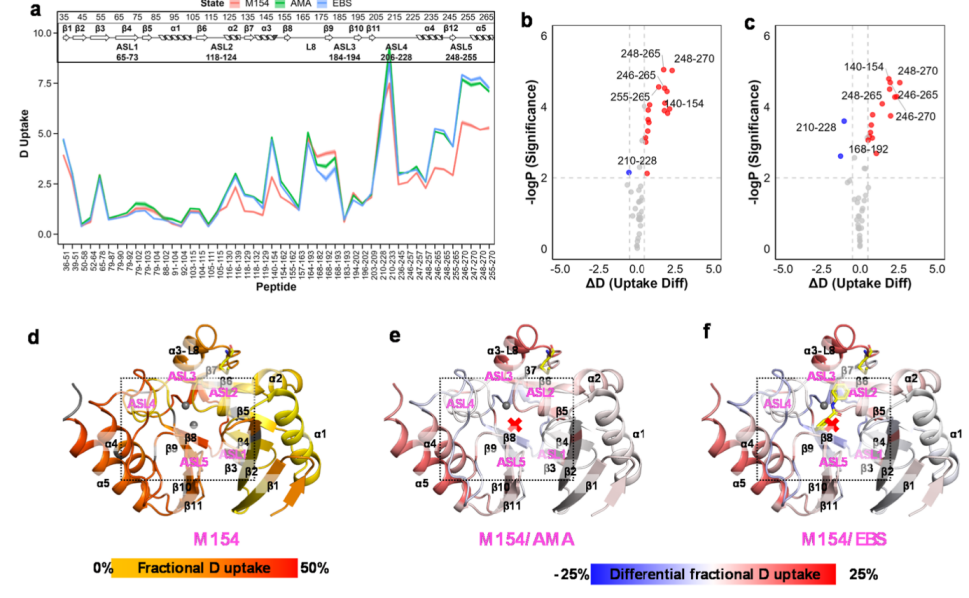
Figure 3
4. Single Mutation M154L Alters the Conformational Dynamics of NDM-1
Further investigation into the α3-L8 region revealed that the M154L mutation accounts for over 50% of the identified NDM variants. Additionally, the M154L mutation significantly enhances the thermal stability of the enzyme, with a decrease in melting temperature (Tm) by 4°C, without affecting its hydrolytic activity. Therefore, we studied how M154L alters conformational stability without loss of activity. The deuterium uptake curves for M154 and L154 are shown in Figure 4; the M154L mutation resulted in an overall reduction in deuterium uptake, indicating that the protein is overall more stable, consistent with its enhanced thermal stability. Furthermore, the mutation significantly increased the flexibility of the α3–L8 region, which may aid the enzyme’s adaptability under zinc-limited conditions. The dynamics of ASL1 and ASL2 between the two structures were roughly equal, while the dynamics of the L8-β9 region and ASL4 moderately decreased after the mutation (Figure 4b). This result is consistent with the reduced turnover rate, which can explain the decline in dynamics after the mutation, as the flexibility of ASL4 is a determinant for substrate binding and release; however, little is known about the role of conformational changes in the L8-β9 region. The authors speculate that this loop region may be involved in the transmission of the allosteric effect induced by the M154L mutation.
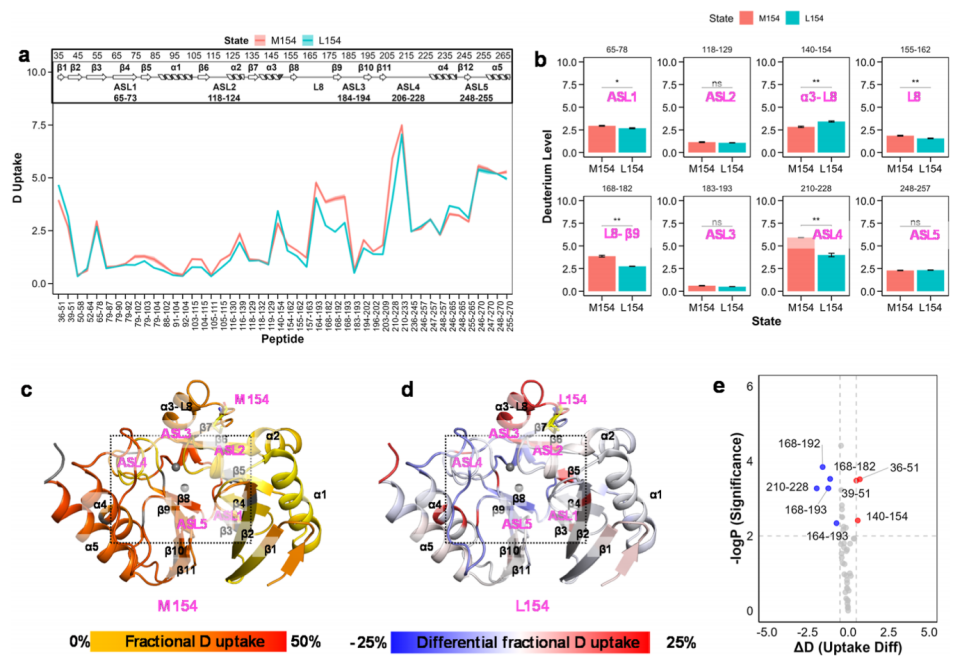
Figure 4
5. Response Mechanism of NDM-1 M154L Mutant to Inhibitors
The authors found that the inhibition by AMA and ebselen depends on the zinc-binding state of NDM. Under zinc-free conditions, the enzyme activity of NDMs significantly decreased in the presence of AMA and ebselen, while under zinc-sufficient conditions, the enzyme activity of NDMs remained unaffected. This result indicates that the inhibition by AMA and ebselen depends on the zinc-binding state of NDM; interestingly, the potency of L/D-captopril does not rely on the zinc scarcity of NDM-1. To confirm the speculations regarding the study of NDM-1, the authors examined the HDX profiles (Figure 5). The uptake profiles of NDM-M154L in the presence of captopril are shown in Figures 5A–C. The M154L variant responded similarly to L/D-captopril as the wild-type, indicating that this mutation does not affect the binding of zinc intercalator-type inhibitors.
Additionally, the study found profound changes in deuterium variations induced by zinc stripping after the M154L mutation, as shown in Figures 5d-f. In vitro activity assays indicated that the M154L mutant maintained higher enzyme activity than the wild-type in the presence of AMA, suggesting that the M154L mutation confers resistance to AMA. The M154L variant exhibited stronger structural protection under zinc-stripped conditions, particularly in the α3–L8 and ASL3 regions, which is consistent with its ability to maintain activity under zinc-limited conditions. The L8-β9 region is the only region that exhibited a similar trend in protection levels between the two variants. The peptide from residues 168-182 in this region was more protected during the M154L mutation (Figure 4b), and subsequently, significantly more protection was observed after AMA and ebselen zinc stripping,highlighting the critical role of the α3-L8-β9 region in regulating the conformational dynamics of NDM under zinc-limited conditions.
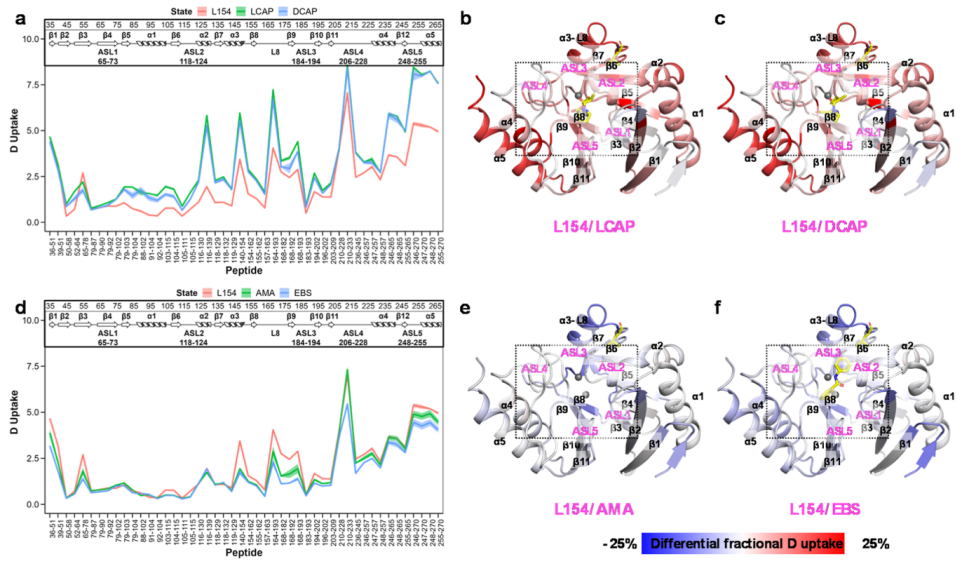
Figure 5
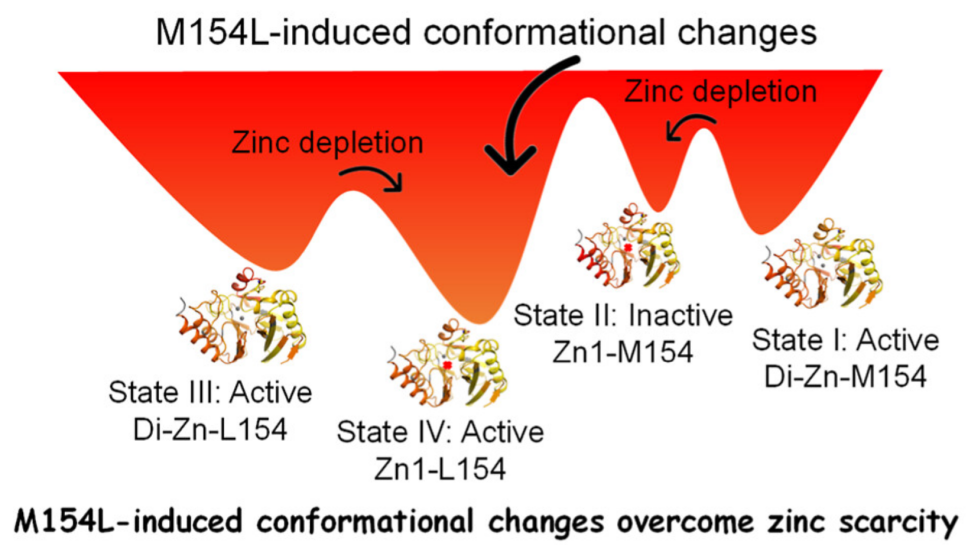
Revealing Pathways of Conformational Rearrangement and Antibiotic Resistance
This work comprehensively studied the inhibitory effects of four different inhibitors on NDM, which were grouped according to three mechanisms of action. Subsequent studies indicated significant conformational changes in the loop regions, particularly the α3-L8-β9 region, where the M154L mutant accounts for 50% of the mutations in this region, followed by an examination of the mutation-induced kinetic properties. To further explore the molecular mechanisms of the widely recognized stability under zinc-limited conditions, the authors systematically validated the changes in the wild-type induced by the perturbation of inhibitors. Using the HDX-MS method, the authors comprehensively studied ten different states of NDM, revealing significant differences in uptake across these states. Furthermore, the authors found that the characteristics of NDMs bound to L- and D-captopril were highly similar. The authors described four different states of NDM, highlighting its structural and functional changes under different zinc conditions (Figure 6). In zinc-rich environments, NDM typically exists in its active di-zinc form (State I). The two zinc ions bind to the enzyme, stabilizing its structure and enabling efficient hydrolysis of β-lactam antibiotics. The presence of zinc is crucial for the enzyme’s catalytic activity, as it coordinates with key residues to form the active site. Under zinc scarcity, NDM may lose Zn2 due to reduced binding affinity, transitioning to an inactive mono-zinc form (State II). In this state, the enzyme’s structure becomes less stable, and its catalytic activity significantly decreases.
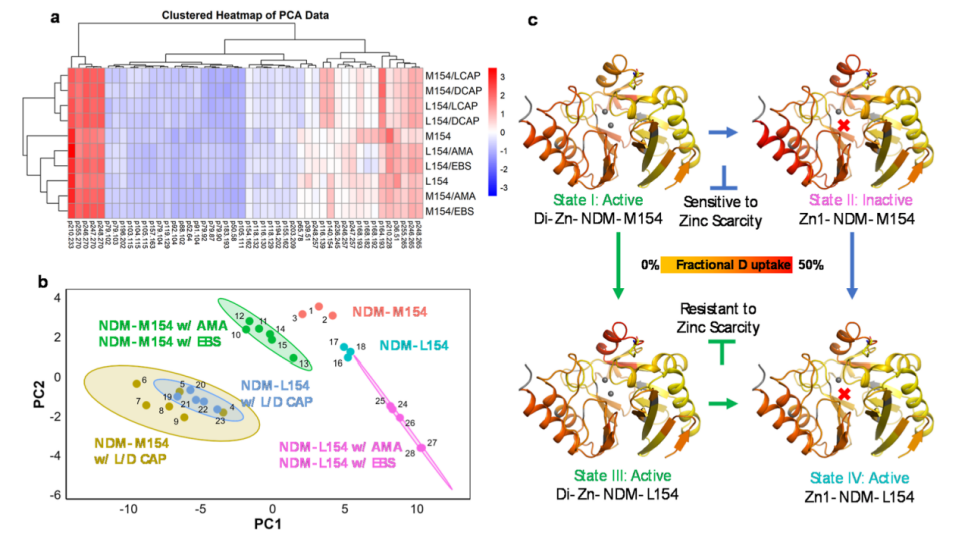
To adapt to zinc scarcity, NDM undergoes evolutionary changes, such as the mutation from M154 to L154 (State III). This mutant enhances zinc-binding affinity, ensuring the enzyme remains active and capable of degrading antibiotics. Under zinc-limited conditions, NDM-L154 can also maintain the structural integrity of its zinc-stripped form (State IV). This adaptability is achieved not only through the structural rearrangement of ASL but also through the regulation of the allosteric region (α3-L8-β9), which stabilizes the enzyme under zinc-limited conditions. The M154L mutation plays a key role in this process, helping to maintain the geometric shape of the enzyme’s active site, allowing it to retain catalytic activity.